從共受體到治療靶點:CD4分子如何調控免疫、影響疾病
日期:2026-03-24 15:11:28
CD4是輔助性T細胞表面的關鍵分子,在適應性免疫應答中發揮核心作用。本文系統梳理了CD4的結構功能、CD4? T細胞亞群分化及其調控機制,重點介紹了TCR、JAK/STAT、TGF-β/SMAD等信號通路在免疫調控中的作用和CD4在HIV/AIDS、自身免疫病、腫瘤等疾病中的研究進展,以及靶向CD4的藥物開發現狀。
1. CD4在免疫系統中的核心地位與研究價值
CD4是適應性免疫系統中的關鍵跨膜糖蛋白,主要表達于輔助性T細胞(CD4+ T細胞)、單核細胞、巨噬細胞和樹突狀細胞表面。其中,在CD4+ T細胞上,CD4作為T細胞受體(TCR)的共受體,通過與主要組織相容性復合體II類(MHC-II)分子結合,協同介導T細胞活化,是適應性免疫應答啟動與調節的重要樞紐。
CD4+ T細胞具有顯著的功能多樣性,可分化為Th1、Th2、Th17和Treg等多個亞群,在免疫輔助、炎癥調控和免疫耐受維持中發揮不可替代的作用 [1]。近年來,隨著成像技術、單細胞分析和計算模型的發展,研究者對CD4分子的認識不斷深化。例如,超分辨成像研究揭示了CD4在靜息T細胞質膜上的納米尺度分布及其受棕櫚酰化影響的特征 [2];腫瘤免疫微環境研究表明,對CD4+ T細胞的自動識別和分類有助于預后評估與治療反應分析 [3];人工智能驅動的成像分析也顯示出區分CD4和CD8細胞的潛力 [4]。
與此同時,針對CD4的藥物開發也不斷推進。以人源化抗CD4單克隆抗體Zanolimumab為例,其可通過抑制TCR信號轉導、誘導Fc依賴性效應機制及下調細胞表面CD4表達等方式抑制CD4+ T細胞,目前正用于皮膚和淋巴結T細胞淋巴瘤相關研究 [5]。這些進展表明,CD4不僅是理解免疫調控機制的關鍵分子,也具有重要的疾病研究與藥物開發價值。
2. CD4的分子特性與基礎功能
2.1 CD4的結構與細胞表面表達
CD4分子通常為單鏈跨膜糖蛋白,由四個免疫球蛋白樣胞外結構域(D1-D4)、一個跨膜區及較短的胞內區組成。其胞外D1和D2結構域主要參與與MHC II類分子的結合,而胞內區則與Lck酪氨酸激酶密切關聯,從而參與TCR信號啟動與調節。
除經典的CD4+ T細胞外,CD4還表達于巨噬細胞、樹突狀細胞和單核細胞等免疫細胞表面,因此其功能并不限于T細胞活化,也與抗原提呈、病毒感染及免疫微環境調控等過程密切相關。近年來,基于TIRF和SOFI的定量成像方法提高了對CD4膜表面納米尺度分布的解析能力,進一步證實了棕櫚酰化對CD4分布模式的重要影響 [2]。此外,人工智能輔助的無標記成像技術也提示,CD4作為細胞表面標志物在細胞識別與分類中具有較高應用潛力 [4]。
2.2 CD4作為TCR共受體的作用機制
在T細胞介導的免疫應答中,CD4的核心作用是協助TCR識別由抗原提呈細胞表面MHC II類分子呈遞的抗原肽。CD4通過與MHC II結合增強TCR-pMHC復合物的穩定性,從而提升T細胞抗原識別的效率與特異性。
除穩定識別復合物外,CD4還可通過胞內區招募并調節p56lck活性。活化后的p56lck可磷酸化TCR復合體中CD3鏈的ITAM基序,進而啟動T細胞早期信號級聯反應。值得注意的是,針對CD4的抗體研究提示,CD4不僅參與激活,也與抑制性調控有關。研究顯示,Zanolimumab可快速抑制早期TCR信號,同時激活CD4相關的p56lck,這提示其可能通過改變p56lck與TCR復合體的耦聯狀態,轉而激活Dok-1、SHIP-1等抑制性信號分子,從而實現對T細胞功能的抑制 [5]。這說明,CD4不僅是簡單的共受體,也是連接識別、激酶招募和信號調控的重要節點。
3. CD4+ T細胞生物學:亞群、分化與調控機制
3.1 CD4+ T細胞亞群及其功能多樣性
CD4+ T細胞具有高度異質性,可根據轉錄因子表達和細胞因子分泌譜分化為不同亞群。經典亞群包括Th1、Th2、Th17和Treg:
- Th1細胞主要分泌IFN-γ,介導抗細胞內病原體和抗腫瘤免疫 [6]
- Th2細胞以IL-4、IL-5和IL-13為特征,主要參與抗寄生蟲免疫和過敏反應
- Th17細胞分泌IL-17A和IL-17F,在抗細胞外病原體感染中發揮作用,但過度活化可促進自身免疫性炎癥 [7]
- Treg細胞則通過Foxp3、IL-10和TGF-β維持免疫耐受并抑制過度免疫反應 [8]
除了經典Th亞群外,Tfh和CD4 CTL也是近年來受到高度關注的重要類型:
- Tfh細胞在B細胞活化、抗體親和力成熟和生發center反應中具有關鍵作用,在慢性感染和慢性移植物抗宿主病中均表現出重要意義 [11-12]
- CD4 CTL則突破了傳統"輔助性T細胞"的概念,具備穿孔素和顆粒酶介導的直接細胞毒性功能,可參與感染細胞或腫瘤細胞清除 [1]
- 此外,一些研究還識別出具有干細胞樣特征的CD4+ T細胞亞群,其可持續補充效應細胞,在慢性炎癥維持中發揮類似"疾病干細胞"的作用 [14]
3.2 CD4+ T細胞分化與可塑性的調控
CD4+ T細胞的分化受抗原強度、細胞因子環境和轉錄因子網絡共同調控:
- 抗原親和力與劑量是決定初始T細胞命運的重要早期信號 [15]
- 外源和內源細胞因子的種類、濃度及比例,則決定分化是呈現突變式還是漸進式變化 [16]
- 例如,IL-2可調節Foxp3 expression,從而影響誘導型Treg的形成 [17]
在細胞內調控層面,Bcl6、Maf、ThPOK、Runx3、Eomes等轉錄因子參與塑造不同CD4+ T細胞亞群的命運與功能 [13], [18-19]。這些因子相互拮抗或協同,使CD4+ T細胞能夠根據感染、炎癥或腫瘤等不同微環境進行動態重塑,從而體現出較強的可塑性。這種可塑性既是免疫應答靈活性的基礎,也增加了疾病研究和治療干預的復雜性。
3.3 CD4+ T細胞的增殖、存活與代謝重編程
CD4+ T細胞在活化后需要經歷快速增殖,并通過代謝重編程滿足效應功能需求。其增殖與存活受IL-7等細胞因子支持,同時也受代謝途徑、線粒體狀態及細胞內能量平衡調控 [27]。不同亞群在代謝模式上存在差異,例如炎癥性效應T細胞通常偏向糖酵解,而調節性T細胞則更多依賴氧化代謝。代謝狀態不僅影響細胞生存,也決定其分化方向和功能輸出,因此代謝調控已成為理解CD4+ T細胞穩態與疾病關聯的重要切入點。
4. 調控CD4+ T細胞功能的關鍵信號通路
4.1 TCR近端信號與鈣調神經磷酸酶-NFAT通路
TCR近端信號是CD4+ T細胞活化的起點。CD4通過協助MHC II識別并招募Lck,促進CD3鏈ITAM磷酸化,進而激活ZAP-70等下游信號分子。隨后,鈣離子信號與鈣調神經磷酸酶-NFAT通路被激活,驅動效應基因表達并調節T細胞分化與功能輸出。該通路對T細胞早期命運決定具有基礎性作用。
4.2 JAK/STAT信號通路
JAK/STAT通路是細胞因子調控CD4+ T細胞分化的核心通路之一。不同細胞因子可通過激活不同STAT成員,促進Th1、Th17或Treg等亞群形成。相關研究表明,某些天然產物或免疫調節因子可通過抑制STAT1、STAT3 or STAT5活性,改變Th1/Th17與Treg之間的平衡,從而在炎癥和自身免疫性疾病中發揮治療潛力 [9-10]。
4.3 TGF-β/SMAD與Notch信號通路
TGF-β/SMAD通路在Treg形成、免疫耐受維持及炎癥調控中具有重要地位;Notch信號則在T細胞發育、分化和功能塑造中發揮輔助調節作用。這兩類通路并非孤立存在,而是與JAK/STAT等通路交互作用,共同參與CD4+ T細胞命運調控。
4.4 cGAS/STING通路與免疫代謝關聯
cGAS/STING通路最初主要被認為參與胞質DNA感知與先天免疫激活,但近年來研究提示,該通路同樣會影響腫瘤免疫環境和T細胞功能狀態。代謝應激、線粒體DNA釋放等事件可激活該通路,進而誘導I型干擾素反應,并影響腫瘤浸潤免疫細胞組成及CD4+ T細胞功能 [29]。
4.5 cAMP及其他相關通路
cAMP信號在CD4+ T細胞免疫調控中同樣具有重要意義。相關研究顯示,Forskolin等cAMP通路激活劑能夠調節自身免疫性炎癥中的CD4+ T cell反應,并與中樞神經系統炎癥緩解相關 [32]。此外,多條信號通路之間存在廣泛串擾,共同決定CD4+ T cell在不同疾病背景下的功能狀態 [28]。
5. CD4+ T細胞在疾病發生發展中的作用
5.1 HIV/AIDS發病機制
HIV感染最典型地體現了CD4在疾病中的核心地位。CD4既是免疫功能評估的重要指標,也是HIV入侵宿主細胞的重要相關分子。CD4+ T cell數量下降直接反映免疫系統受損程度,而病毒載量、宿主個體差異及免疫重建能力等因素共同決定疾病進展速度。因此,CD4計數長期以來一直是HIV感染管理與療效評估的重要參數。
5.2 自身免疫性疾病與炎癥
在類風濕關節炎、系統性紅斑狼瘡、實驗性自身免疫性腦脊髓炎等疾病中,CD4+ T cell亞群失衡是關鍵病理機制之一。尤其是Th17/Treg失衡,常與慢性炎癥放大和組織損傷相關 [9-10]。多種代謝干預、細胞療法及信號通路調節策略,均顯示出通過重塑CD4+ T cell功能來緩解疾病的潛力 [26], [30]。
5.3 癌癥免疫與免疫治療
在腫瘤免疫中,CD4+ T cell既能輔助CD8+ T cell發揮殺傷作用,也可獨立介導抗腫瘤免疫,甚至作為療效預測生物標志物。這說明,CD4+ T cell不僅是腫瘤免疫中的"輔助者",在一定條件下也可能是關鍵"執行者"。
5.4 其他感染性疾病
除HIV外,CD4+ T cell在流感等急慢性感染中同樣發揮重要作用。特別是CD4 CTL的發現,提示CD4+ T cell在抗病毒過程中不僅負責調控,也可直接參與感染細胞清除 [1]。這拓展了對CD4+ T cell在感染免疫中功能邊界的理解。
6. CD4靶向藥物最新研究進展
目前,靶向CD4的藥物類型多樣,涵蓋單抗、雙抗、CAR-T及小分子藥物;在研適應癥以HIV感染為主,并拓展至特應性皮炎、腫瘤等領域;研發階段覆蓋已上市品種至臨床早期,部分在研管線列舉如下。
| 藥物 | 靶點 | 藥物類型 | 在研適應癥 | 在研機構 | 最高研發階段 |
|---|---|---|---|---|---|
| 艾巴利珠單抗 | CD4 x Viral fusion proteins | 雙特異性抗體 | HIV感染 | Theratechnologies, Inc. | 批準上市 |
| Semzuvolimab | CD4 | 單克隆抗體 | HIV感染 | 聯生藥大中華控股有限公司 | 聯合生物制藥股份有限公司 | 聯亞藥業股份有限公司 | 臨床3期 |
| Mosedipimod | CD4 x CD8 | 小分子化藥 | 中度特應性皮炎 | 重度特應性皮炎 | 代謝功能障礙相關脂肪性肝炎 | ENZYCHEM LIFESCIENCES Corp. | 臨床2期 |
| Iotivibart | CD4 x HIV envelope protein gp120 | 單克隆抗體 | HIV感染 | ViiV Healthcare UK Ltd. | ViiV Healthcare Ltd. | 臨床2期 |
| TMB-365 | CD4 | 單克隆抗體 | HIV感染 | 中裕新藥股份有限公司 | 臨床2期 |
| IB-MS | CD4 x CD8 | 小分子化藥 | 多發性硬化癥 | INNOBIOSCIENCE LLC | 臨床2期 |
| CD4 CAR T cell therapy (iCell Gene Therapeutics) | CD4 | 自體CAR-T | 慢性粒單核細胞白血病 | T細胞白血病 | T細胞淋巴瘤 | iCell Gene Therapeutics, Inc. | 臨床1期 |
| IT-1208(Kyowa Hakko Kirin Pharma, Inc.) | CD4 | 單克隆抗體 | 腫瘤 | IDAC Theranostics, Inc. | Ono Pharmaceutical Co., Ltd. | Kyowa Kirin Co., Ltd. | 臨床1期 |
| Autologous CD4 CAR T-cells(Indiana University) | CD4 | 自體CAR-T | 難治性急性髓細胞白血病 | 復發性急性髓細胞白血病 | iCell Gene Therapeutics, Inc. | 臨床1期 |
| VRCHIVMAB0115-00-AB | CD4 | 單克隆抗體 | HIV感染 | National Institute of Allergy & Infectious Diseases | 臨床1期 |
| BG-8962 | CD4 | 生物藥 | HIV感染 | National Institute of Allergy & Infectious Diseases | 臨床1期 |
| LM49 | CD4 | 小分子化藥 | 糖尿病腎病 | 山西醫科大學 | 山西中醫藥大學 | 臨床1期 |
(數據截止到2026年3月16日,來源于synapse)
7. CD4研究工具
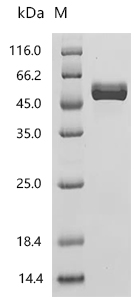
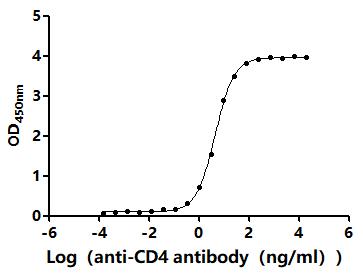
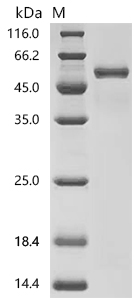
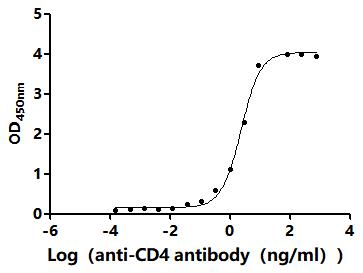
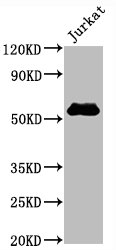
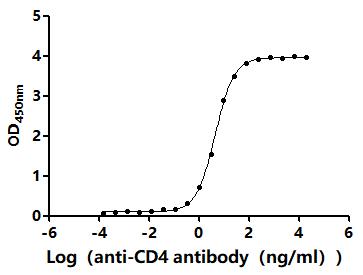
CD4不僅是輔助性T細胞的重要表面分子,更是連接抗原識別、信號轉導、細胞分化和疾病進程的關鍵樞紐。圍繞CD4及CD4+ T cell的研究,已經從傳統的"輔助功能"拓展到細胞毒性、代謝調控、組織微環境適應和疾病持續化機制等多個層面。華美生物提供CD4重組蛋白、抗體及ELISA試劑盒產品,助力您進行相關機制研究及靶向藥物開發。
[1] Deborah M. Brown, Anna T. Lampe, Aspen M. Workman. (2016). The Differentiation and Protective Function of Cytolytic CD4 T Cells in Influenza Infection.
[2] Tomas Lukes, Daniela Glatzova, Zuzana Kvicalova, Florian Levet, Ales Benda, Tomas Brdicka, Theo Lasser, Marek Cebecauer. (2017). Quantifying protein densities on cell membranes using super-resolution optical fluctuation imaging.
[3] Mika?l Simard, Zhuoyan Shen, Konstantin Br?utigam, Rasha Abu-Eid, Maria A. Hawkins, Charles-Antoine Collins-Fekete. (2024). Immunocto: a massive immune cell database auto-generated for histopathology.
[4] Xin Shu, Sameera Sansre, Di Jin, Xiangxiang Zeng, Kai-Yu Tong, Rishikesh Pandey, Renjie Zhou. (2020). Artificial Intelligence Enabled Reagent-free Imaging Hematology Analyzer.
[5] D. Rider, C. Havenith, R. De Ridder, J. Schuurman, C. Favre, J. Cooper, S. Walker, O. Baadsgaard, S. Marschner, Jan G J vandeWinkel, J. Cambier, P. Parren, D. Alexander. (2007). A human CD4 monoclonal antibody for the treatment of T-cell lymphoma combines inhibition of T-cell signaling by a dual mechanism with potent Fc-dependent effector activity.
[6] J. T. Ulrich-Lewis, K. Draves, Kelsey Roe, M. O'Connor, E. Clark, D. Fuller. (2022). STING Is Required in Conventional Dendritic Cells for DNA Vaccine Induction of Type I T Helper Cell- Dependent Antibody Responses.
[7] Constance Renault, N. Veyrenche, F. Mennechet, A. Bedin, J. Routy, P. Van de Perre, J. Reynes, E. Tuaillon. (2022). Th17 CD4+ T-Cell as a Preferential Target for HIV Reservoirs.
[8] Rong Wang, Keliang Huang. (2020). CCL11 increases the proportion of CD4+CD25+Foxp3+ Treg cells and the production of IL-2 and TGF-β by CD4+ T cells via the STAT5 signaling pathway.
[9] Jiaxu Qin, Xiangwei Zheng, Yancheng He, Yang Hong, Shuang Liang, X. Fang. (2022). The regulation of T helper cell polarization by the diterpenoid fraction of Rhododendron molle based on the JAK/STAT signaling pathway.
[10] Fanping Wang, Yonghui Yang, Zhixin Li, Yan Wang, Zhenchao Zhang, Wei Zhang, Yonghui Mu, Jing-ke Yang, Lili Yu, Mingyong Wang. (2022). Mannan-Binding Lectin Regulates the Th17/Treg Axis Through JAK/STAT and TGF-β/SMAD Signaling Against Candida albicans Infection.
[11] R. Deng, C. Hurtz, Qingxiao Song, Chanyu Yue, G. Xiao, Hua Yu, Xiwei Wu, M. Muschen, S. Forman, P. Martin, D. Zeng. (2017). Extrafollicular CD4+ T-B interactions are sufficient for inducing autoimmune-like chronic graft-versus-host disease.
[12] Laura M. Fahey, Elizabeth B. Wilson, Heidi J. Elsaesser, Chris Fistonich, D. McGavern, D. Brooks. (2011). Viral persistence redirects CD4 T cell differentiation toward T follicular helper cells.
[13] A. Dejean, Emeline Joulia, T. Walzer. (2018). The role of Eomes in human CD4 T cell differentiation: A question of context.
[14] Yuki Sato, Abhinav Jain, Shozo Ohtsuki, Hirohisa Okuyama, Ines Sturmlechner, Yoshinori Takashima, Kevin-Phu C. Le, Melanie C. Bois, Gerald J. Berry, K. Warrington, J. Goronzy, C. Weyand. (2023). Stem-like CD4+ T cells in perivascular tertiary lymphoid structures sustain autoimmune vasculitis.
[15] S. Keck, M. Schmaler, S. Ganter, Lena Wyss, Susanne G. Oberle, E. Huseby, D. Zehn, C. King. (2014). Antigen affinity and antigen dose exert distinct influences on CD4 T-cell differentiation.
[16] M. Martínez-Sánchez, L. Huerta, E. álvarez-Buylla, Carlos Villarreal Luján. (2018). Role of Cytokine Combinations on CD4+ T Cell Differentiation, Partial Polarization, and Plasticity: Continuous Network Modeling Approach.
[17] E. Zorn, E. Nelson, M. Mohseni, F. Porcheray, Haesook T. Kim, Despina Litsa, R. Bellucci, E. Raderschall, C. Canning, R. Soiffer, D. Frank, J. Ritz. (2006). IL-2 regulates FOXP3 expression in human CD4+CD25+ regulatory T cells through a STAT-dependent mechanism and induces the expansion of these cells in vivo.
[18] M. Kroenke, D. Eto, M. Locci, Michael Cho, T. Davidson, E. Haddad, S. Crotty. (2012). Bcl6 and Maf Cooperate To Instruct Human Follicular Helper CD4 T Cell Differentiation.
[19] Megan A. Luckey, M. Kimura, A. Waickman, L. Feigenbaum, A. Singer, Jung-Hyun Park. (2014). The transcription factor ThPOK suppresses Runx3 and imposes CD4+ lineage fate by inducing the SOCS suppressors of cytokine signaling.
[20] Qingtian Li, Jia Zou, Mingjun Wang, Xilai Ding, I. Chepelev, Xikun Zhou, Wei Zhao, Gang Wei, J. Cui, K. Zhao, Helen Y Wang, Rong-Fu Wang. (2014). Critical role of histone demethylase Jmjd3 in the regulation of CD4+ T-cell differentiation.
[21] Y. Shu, Qinghua Hu, H. Long, Christopher Chang, Q. Lu, R. Xiao. (2017). Epigenetic Variability of CD4+CD25+ Tregs Contributes to the Pathogenesis of Autoimmune Diseases.
[22] Sarah A. LaMere, Ryan C. Thompson, Xiangzhi Meng, H. Komori, Adam M. Mark, D. Salomon. (2017). H3K27 Methylation Dynamics during CD4 T Cell Activation: Regulation of JAK/STAT and IL12RB2 Expression by JMJD3.
[23] M. Martínez-Sánchez, L. Mendoza, C. Villarreal, E. álvarez-Buylla. (2015). A Minimal Regulatory Network of Extrinsic and Intrinsic Factors Recovers Observed Patterns of CD4+ T Cell Differentiation and Plasticity.
[24] B. L. Puniya, R. G. Todd, Akram Mohammed, Deborah M. Brown, Matteo Barberis, T. Helikar. (2018). A Mechanistic Computational Model Reveals That Plasticity of CD4+ T Cell Differentiation Is a Function of Cytokine Composition and Dosage.
[25] V. Proserpio, A. Piccolo, Liora Haim-Vilmovsky, G. Kar, T. L?nnberg, Valentine Svensson, J. Pramanik, K. Natarajan, Weichao Zhai, Xiuwei Zhang, G. Donati, M. Kayikci, J. Kotar, A. McKenzie, Ruddy Montandon, O. Billker, S. Woodhouse, P. Cicuta, M. Nicodemi, S. Teichmann. (2016). Single-cell analysis of CD4+ T-cell differentiation reveals three major cell states and progressive acceleration of proliferation.
[26] S. Tullius, H. Biefer, Suyan Li, A. Trachtenberg, K. Edtinger, M. Quante, Felix Krenzien, H. Uehara, Xiaoyong Yang, H. Kissick, W. Kuo, I. Ghiran, M. A. de la Fuente, M. Arredouani, Virginia Camacho, J. Tigges, Vasilis Toxavidis, Rachid El Fatimy, Brian D. Smith, A. Vasudevan, A. Elkhal. (2014). NAD+ protects against EAE by regulating CD4+ T-cell differentiation.
[27] Nizar Chetoui, M. Boisvert, S. Gendron, F. Aoudjit. (2010). Interleukin‐7 promotes the survival of human CD4+ effector/memory T cells by up‐regulating Bcl‐2 proteins and activating the JAK/STAT signalling pathway.
[28] M. S. Alam, Y. Maekawa, A. Kitamura, K. Tanigaki, T. Yoshimoto, K. Kishihara, K. Yasutomo. (2010). Notch signaling drives IL-22 secretion in CD4+ T cells by stimulating the aryl hydrocarbon receptor.
[29] Shengpan Wu, Baojun Wang, Hong-zhao Li, Hanfeng Wang, Songliang Du, Xing Huang, Yang Fan, Yu Gao, L. Gu, Qingbo Huang, Jianjun Chen, Xu Zhang, Yan Huang, Xin Ma. (2024). Targeting STING elicits GSDMD-dependent pyroptosis and boosts anti-tumor immunity in renal cell carcinoma.
[30] Dongmei Li, Idalia Cruz, Yahui Feng, Maha Moussa, Jie Cheng, Digvijay Patil, Alexander Kroemer, Joseph A. Bellanti. (2025). Divergent Immunomodulatory Roles of Fungal DNA in Shaping Treg and Inflammatory Responses.
[31] C. Elsner, Aparna Ponnurangam, J. Kazmierski, T. Zillinger, Jenny Jansen, D. Todt, K. D?hner, Shuting Xu, A. Ducroux, Nils Kriedemann, Angelina Malassa, P. Larsen, G. Hartmann, W. Barchet, E. Steinmann, U. Kalinke, B. Sodeik, C. Goffinet. (2020). Absence of cGAS-mediated type I IFN responses in HIV-1--infected T cells.
[32] Tatyana Veremeyko, Amanda W. Y. Yung, M. Dukhinova, Inna S. Kuznetsova, I. Pomytkin, A. Lyundup, T. Strekalova, N. Barteneva, E. Ponomarev. (2018). Cyclic AMP Pathway Suppress Autoimmune Neuroinflammation by Inhibiting Functions of Encephalitogenic CD4 T Cells and Enhancing M2 Macrophage Polarization at the Site of Inflammation.















