CCR4:免疫治療與炎癥干預的核心靶點
日期:2026-03-17 08:53:03
1. CCR4靶點概述:生理功能與病理調控的核心作用
C-C 趨化因子受體 4(CCR4)是 G 蛋白偶聯受體家族的重要成員,主要參與免疫細胞的募集、遷移及功能調控。作為 Th2 細胞和調節性 T 細胞(Regulatory T cells, Treg)的關鍵表面受體之一,CCR4 通過識別其配體 CCL17(TARC)和 CCL22(MDC),在免疫穩態維持、炎癥反應及免疫耐受形成過程中發揮重要作用。近年來的研究表明,CCR4 的異常表達或功能失衡與多種疾病密切相關,尤其在血液系統惡性腫瘤、實體瘤、自身免疫性疾病及慢性炎癥性疾病中表現突出。CCR4 不僅通過募集免疫抑制性 Treg 改變局部免疫微環境,還可直接參與腫瘤細胞遷移、侵襲及耐藥調控。隨著 CCR4 靶向抗體、小分子拮抗劑及細胞治療策略的不斷推進,該受體已成為免疫治療和炎癥干預領域的重要研究靶點。
2. CCR4的分子結構與配體互作機制
CCR4 是一種典型的七次跨膜 G 蛋白偶聯受體,主要表達于 Th2、Treg、樹突狀細胞、巨噬細胞及細胞因子誘導殺傷細胞等多種免疫細胞表面 [4–7]。在健康個體中,CCR4 常被視為 Th2、Th17 和 Treg 細胞的重要標志物 [4],而在多種腫瘤細胞中,其異常表達與不良預后密切相關 [8–10]。CCR4 的主要配體為 CCL17(TARC)和 CCL22(MDC),二者由樹突狀細胞和巨噬細胞等多種細胞產生,通過募集 CCR4 陽性免疫細胞在炎癥及腫瘤微環境中發揮重要調控作用 [6–8]。在腫瘤微環境中,M2 型巨噬細胞可顯著增強 CCL17 的分泌,進而促進腫瘤細胞的遷移、侵襲及干性維持 [8]。配體-受體結合后,CCR4 可激活多條信號通路,并在不同細胞類型和疾病背景下呈現出顯著的功能多樣性。
3. CCR4信號通路解析:免疫調控與疾病進展的分子基礎
3.1 G 蛋白依賴性信號通路
CCR4 主要通過 Gαi 蛋白介導下游信號轉導。配體結合后,Gβγ 亞基可激活 Rho 激酶和 p38 MAPK 通路,調控細胞骨架重排、趨化遷移及炎癥反應 [13]。此外,在不同病理環境中,CCR4 還可通過 PI3K/AKT 等信號通路影響細胞存活、代謝及耐藥性 [11,12]。
3.2 G 蛋白非依賴性信號及偏向性激動
β-arrestin2 在 CCR4 信號轉導中具有重要作用,既參與受體內吞,又可介導獨立的信號級聯反應。在過敏性炎癥模型中,CCL22-CCR4-β-arrestin2 軸被證實在 Th2 細胞趨化和炎癥加重中發揮關鍵作用 [13]。不同配體在特定細胞環境下可能呈現偏向性信號激活模式,為靶向干預提供了新的思路 [14]。
3.3 CCR4 在免疫細胞遷移與功能中的作用
CCR4 是 Th2 細胞和 Treg 細胞趨化的重要調控因子。在腫瘤微環境中,CCR4? Treg 的富集與免疫抑制和不良預后密切相關 [15]。此外,CCR4 還參與 CIK 細胞、樹突狀細胞及巨噬細胞功能調控,在抗腫瘤免疫、炎癥反應及免疫耐受形成中呈現雙重作用。
3.4 CCR4 調控的其他生物學過程
CCR4 通過 ERK/AKT、EZH2-DNMT3A 等多條通路調控腫瘤細胞遷移、侵襲、上皮-間質轉化及耐藥過程 [3,8,16],進一步凸顯其在腫瘤進展中的核心地位。
3.5 CCR4 與其他趨化因子受體的協同與差異
在胸腺發育和中樞耐受建立過程中,CCR4 與 CCR7 在不同階段和細胞亞群中發揮互補作用,共同確保自身反應性 T 細胞的有效清除 [17]。
4. CCR4在多系統疾病中的致病機制:腫瘤、自身免疫病及其他適應癥
4.1 血液系統惡性腫瘤
CCR4 在多種 T 細胞來源的血液系統惡性腫瘤中高度表達,尤其在成人 T 細胞白血病/淋巴瘤(ATL)和皮膚 T 細胞淋巴瘤(CTCL)中具有明確的病理學意義。CCR4? 腫瘤細胞可通過其配體 CCL17 和 CCL22 實現定向遷移和組織浸潤,同時逃避免疫監視。臨床研究顯示,CCR4 的高表達通常與疾病進展、侵襲性增強及不良預后密切相關。基于這一特征開發的人源化抗 CCR4 單克隆抗體 Mogamulizumab,通過抗體依賴的細胞介導細胞毒作用(ADCC)有效清除 CCR4? 腫瘤細胞,并可同時耗竭免疫抑制性的 Treg,從而改善抗腫瘤免疫反應 [2,18]。該藥物已獲批用于復發或難治性 CTCL 和外周 T 細胞淋巴瘤,驗證了 CCR4 作為治療靶點的臨床可行性。
4.2 實體瘤
除血液系統腫瘤外,CCR4 在多種實體瘤中同樣發揮重要作用,包括肝細胞癌、肺腺癌、乳腺癌及結直腸癌等。研究發現,腫瘤相關巨噬細胞和樹突狀細胞可分泌大量 CCL17 和 CCL22,選擇性募集 CCR4? Treg 至腫瘤微環境,導致局部免疫抑制并削弱抗腫瘤免疫應答 [8,15]。此外,部分實體瘤細胞本身亦表達 CCR4,其激活可通過 PI3K/AKT、ERK 等信號通路促進腫瘤細胞增殖、遷移和上皮-間質轉化,從而增強侵襲性和轉移潛能 [3,8]。這些發現為 CCR4 靶向治療在實體瘤中的應用提供了理論基礎,并推動了 CCR4 拮抗劑與免疫檢查點抑制劑聯合治療策略的探索 [19,20]。
4.3 炎癥性與自身免疫性疾病
CCR4 在炎癥性和自身免疫性疾病中的作用主要與 Th2 和 Treg 細胞遷移密切相關。在類風濕關節炎、特應性皮炎及系統性紅斑狼瘡等疾病中,CCR4-CCL17/CCL22 軸的異常激活可導致免疫細胞在靶組織中過度聚集,從而加重炎癥反應和組織損傷 [1]。在部分疾病背景下,CCR4? Treg 的募集可能具有雙重作用:一方面有助于抑制過度炎癥,另一方面在慢性炎癥狀態下可能促進免疫失衡并延緩組織修復。這種功能的情境依賴性也提示 CCR4 靶向干預需充分考慮疾病階段和免疫狀態。
4.4 代謝相關疾病
近年來,CCR4 在代謝性疾病中的作用逐漸受到關注。在肥胖及其相關并發癥中,CCR4 及其配體與脂肪組織慢性低度炎癥、血管內皮功能障礙密切相關。CCR4 介導的免疫細胞募集可能參與胰島素抵抗和代謝紊亂的發生過程,為炎癥-代謝交叉調控機制提供了新的研究視角。
4.5 病毒感染相關疾病
在 HTLV-1 和 HIV-1 感染中,CCR4? CD4? T 細胞被認為是重要的病毒儲存庫。CCR4 的高表達不僅與病毒持續存在相關,還可能影響免疫重建和疾病進展 [4,21]。這些發現提示,CCR4 在病毒感染相關免疫病理過程中具有潛在的干預價值。
4.6 神經性疼痛
動物實驗研究表明,CCR4 及其配體在神經性疼痛的發生發展中發揮調控作用。CCR4 拮抗劑可顯著緩解疼痛行為,并增強阿片類鎮痛藥物的療效 [22],為慢性疼痛的多靶點聯合治療提供了新的研究方向。
5. CCR4靶向藥物研發進展:已上市藥物與在研管線全景
近年來,靶向CCR4的藥物研發取得了顯著進展,涵蓋了從單克隆抗體、小分子化藥到CAR-T細胞療法等多種藥物類型。其中,莫格利珠單抗作為CCR4拮抗劑的代表藥物,已在菌樣真菌病、塞扎里綜合征及外周T細胞淋巴瘤等適應癥中獲得批準上市。其他在研藥物如Tivumecirnon Besylate、Autologous CCR4 CAR T cells等正處于臨床2期和1期階段,主要針對轉移性胃癌、晚期癌癥及T細胞淋巴瘤等惡性腫瘤。此外,多個臨床前研究項目正在探索CCR4拮抗劑在膿毒癥、肝癌、哮喘、鼻炎、貧血及腦卒中等多種疾病中的潛力。總體來看,CCR4靶向藥物的研發呈現出多樣化的發展趨勢,腫瘤仍是主要研究方向,同時也在向其他疾病領域拓展。
| 藥物 | 作用機制 | 藥物類型 | 在研適應癥 | 在研機構 | 最高研發階段 |
|---|---|---|---|---|---|
| 莫格利珠單抗 | CCR4拮抗劑 | 單克隆抗體 | 蕈樣真菌病 | 塞扎里綜合征 | 外周T細胞淋巴瘤等 | Kyowa Kirin Holdings BV | Kyowa Kirin, Inc. | Kyowa Kirin Australia Pty Ltd. | Kyowa Kirin Co., Ltd. | Kyowa Kirin, Inc. /Sancuso/ | 批準上市 |
| Tivumecirnon Besylate | CCR4拮抗劑 | 小分子化藥 | 轉移性胃癌 | 晚期癌癥 | Rapt Therapeutics, Inc. | Hanmi Pharmaceutical Co., Ltd. | 臨床2期 |
| Autologous CCR4 CAR T cells(NCI) | CCR4 modulators | 自體CAR-T | - | National Cancer Institute | 臨床1期 |
| DB07845 | CCR4拮抗劑 | 小分子化藥 | 膿毒癥 | 嘉興市中醫醫院 | 臨床前 |
| CCR4-351 | CCR4拮抗劑 | 小分子化藥 | 成人T細胞白血病/淋巴瘤 | 腫瘤 | Albert Einstein College of Medicine, Inc. | Rapt Therapeutics, Inc. | 臨床前 |
| NTX-471 | CCR4 modulators | CD47調節劑 | mRNA編碼的雙抗 | 貧血 | Nutcracker Therapeutics, Inc. | 臨床前 |
| CCR4抑制劑(ChemoCentryx) | CCR4拮抗劑 | 小分子化藥 | 哮喘 | 鼻炎 | ChemoCentryx, Inc. | 臨床前 |
| CCR4拮抗劑(Aurigene) | CCR4拮抗劑 | 化學藥 | 腫瘤 | Aurigene Oncology Ltd. | 臨床前 |
| BJY-809 | CCR4拮抗劑 | 小分子化藥 | 肝癌 | 八加一藥業股份有限公司 | 臨床前 |
| Small molecule therapeutic(Collaboration Pharmaceuticals) | CCR3拮抗劑 | CCR4拮抗劑 | 小分子化藥 | 腦卒中 | Collaborations Pharmaceuticals, Inc. | 臨床前 |
| TQB-2619 | CCR4拮抗劑 | 生物藥 | 慢性淋巴細胞白血病 | T細胞淋巴瘤 | 正大天晴藥業集團股份有限公司 | 臨床前 |
(數據截止到2026年3月11日,來源于synapse)
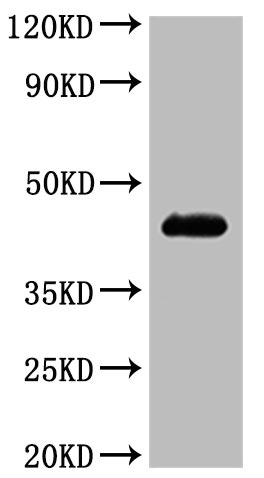
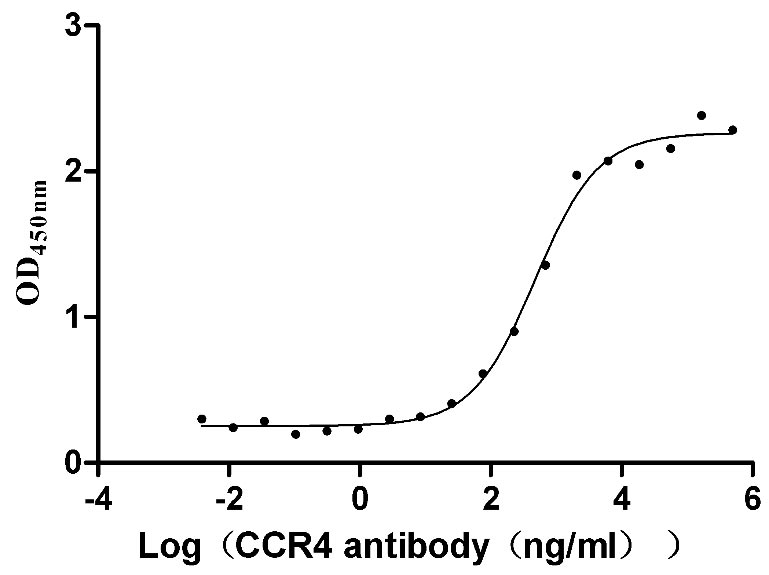
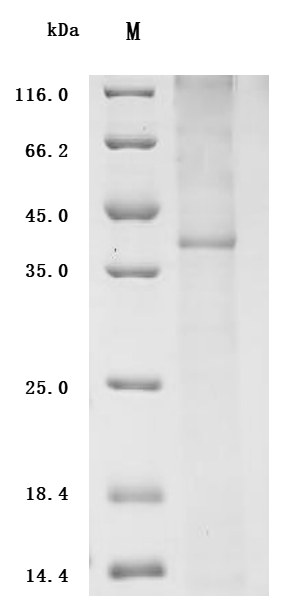
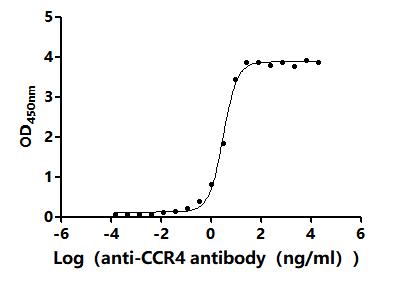
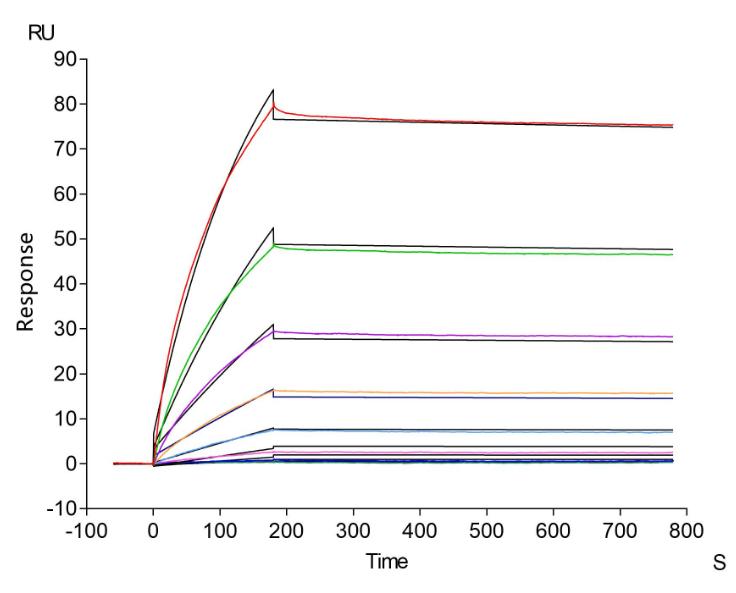
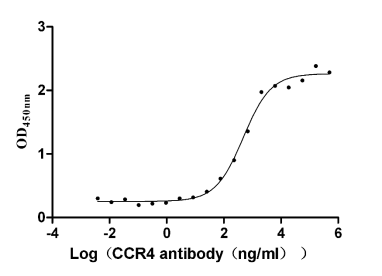
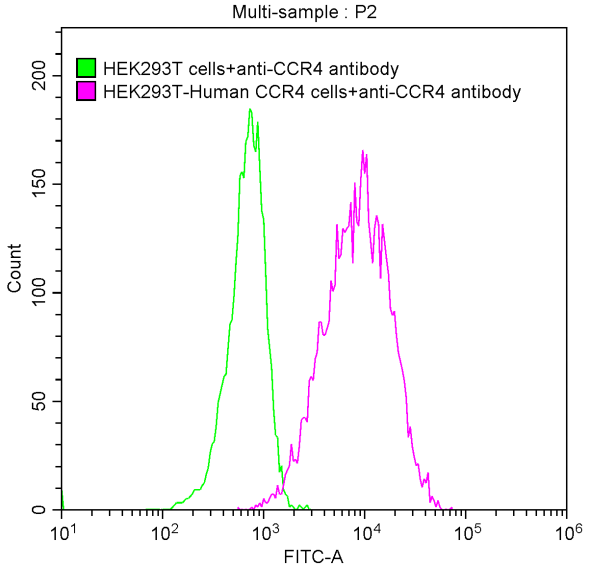
6. CCR4研究工具推薦:重組蛋白與抗體產品選型指南
華美生物提供CCR4重組蛋白、抗體產品,助力您進行相關機制研究及靶向藥物開發。
[1] Kayoko Hase, K. Tani, Teruki Shimizu, Y. Ohmoto, K. Matsushima, S. Sone.(2001). Increased CCR4 expression in active systemic lupus erythematosus.
[2] Kenji Ishitsuka, Tomoharu Yasukawa, Yukie Tsuji.(2024). Safety and effectiveness of mogamulizumab in relapsed or refractory CC chemokine receptor 4‐positive peripheral T‐cell lymphoma and relapsed or refractory cutaneous T‐cell lymphoma: A post‐marketing surveillance in Japan.
[3] Xi Cheng, Huo Wu, Zhi-Jian Jin, Ding Ma, Stanley Yuen, Xiaoqian Jing, Minmin Shi, B. Shen, C. Peng, R. Zhao, Wei-hua Qiu.(2017). Up-regulation of chemokine receptor CCR4 is associated with Human Hepatocellular Carcinoma malignant behavior.
[4] Y. Yamano, N. Araya, Tomoo Sato, A. Utsunomiya, Kazuko Azakami, D. Hasegawa, Toshihiko Izumi, Hidetoshi Fujita, S. Aratani, N. Yagishita, Ryoji Fujii, K. Nishioka, S. Jacobson, T. Nakajima.(2009). Abnormally High Levels of Virus-Infected IFN-γ+CCR4+CD4+CD25+ T Cells in a Retrovirus-Associated Neuroinflammatory Disorder.
[5] C. Carola, Andrew Salazar, C. Rakers, F. Himbert, Q. Do, P. Bernard, Joerg von Hagen.(2021). A Cornflower Extract Containing N-Feruloylserotonin Reduces Inflammation in Human Skin by Neutralizing CCL17 and CCL22 and Inhibiting COX-2 and 5-LOX.
[6] Yongjian Dong, Shuhui Gao, Xue-fang Zhang, Jing Kou, Jing Liu, Ting Ye, Han Shen.(2021). CCL17 and CCL22 induce CCR4 receptor expression and promote cytokine-induced killer cells migration.
[7] Inés Lecoq, K. Kopp, M. Chapellier, Panagiotis Mantas, E. Martinenaite, Maria Perez-Penco, Lars Rønn Olsen, M. Zocca, A. Wakatsuki Pedersen, M. Andersen.(2022). CCL22-based peptide vaccines induce anti-cancer immunity by modulating tumor microenvironment.
[8] Chun Jin, Liangliang Lu, Jian Gao, Ling Chen.(2024). M2-like Macrophages-derived CCL17 Promotes Esophageal Squamous Cell Carcinoma Metastasis and Stemness via Activating CCR4-mediated ERK/PD-L1 Pathway..
[9] A. Maolake, K. Izumi, K. Shigehara, Ariunbold Natsagdorj, H. Iwamoto, S. Kadomoto, Yuta Takezawa, Kazuaki Machioka, K. Narimoto, M. Namiki, Wen-Jye Lin, Guzailinuer Wufuer, A. Mizokami.(2016). Tumor-associated macrophages promote prostate cancer migration through activation of the CCL22–CCR4 axis.
[10] W. Gan, Baoye Sun, Zhangfu Yang, Cheng Ye, Zhutao Wang, Cheng Zhou, Guo-Qiang Sun, Yong Yi, Shuang-Jian Qiu.(2024). Enhancing hepatocellular carcinoma management: prognostic value of integrated CCL17, CCR4, CD73, and HHLA2 expression analysis.
[11] Lichao Zhang, Xiaoqing Lu, Yuanzhi Xu, Xiaoqin La, Jinmiao Tian, Aiping Li, Hanqing Li, Changxin Wu, Yanfeng Xi, Guisheng Song, Zhaocai Zhou, Wenqi Bai, Liwei An, Zhuoyu Li.(2023). Tumor-associated macrophages confer colorectal cancer 5-fluorouracil resistance by promoting MRP1 membrane translocation via an intercellular CXCL17/CXCL22–CCR4–ATF6–GRP78 axis.
[12] Shuixiang Deng, Peng Jin, P. Sherchan, Shengpeng Liu, Yuhui Cui, Lei Huang, John H. Zhang, Ye Gong, Jiping Tang.(2021). Recombinant CCL17-dependent CCR4 activation alleviates neuroinflammation and neuronal apoptosis through the PI3K/AKT/Foxo1 signaling pathway after ICH in mice.
[13] Rui Lin, Y. Choi, David A Zidar, Julia K. L. Walker.(2018). β‐Arrestin‐2‐Dependent Signaling Promotes CCR4‐mediated Chemotaxis of Murine T‐Helper Type 2 Cells.
[14] H. Lim, J. R. Lane, Meritxell Canals, M. Stone.(2021). Systematic Assessment of Chemokine Signaling at Chemokine Receptors CCR4, CCR7 and CCR10.
[15] Tania Sarkar, Subhanki Dhar, D. Chakraborty, Subhadip Pati, S. Bose, A. Panda, Udit Basak, S. Chakraborty, S. Mukherjee, A. Guin, Kuladip Jana, Diptendra K. Sarkar, Gaurisankar Sa.(2022). FOXP3/HAT1 Axis Controls Treg Infiltration in the Tumor Microenvironment by Inducing CCR4 Expression in Breast Cancer.
[16] Li Zhang, Sijuan Tian, Jie Chang, Shimin Quan, Ting Yang, Minyi Zhao, Li Wang, Xiaofeng Yang.(2024). Activation of the CCL22/CCR4 causing EMT process remodeling under EZH2-mediated epigenetic regulation in cervical carcinoma.
[17] Y. Li, J. Srinivasan, Hilary J. Selden, Lauren I. R. Ehrlich.(2023). CCR4 and CCR7 differentially regulate thymocyte localization with distinct outcomes for central tolerance.
[18] Z. Maru?i?, E. Calonje.(2021). Skin adnexal tumours in a tertiary dermatopathology service.
[19] Do-Youn Oh, Min-Hee Ryu, Jun-Eul Hwang, Jaeyong Cho, D. Y. Zang, Sang Cheul Oh, J. Lee, Keun-Wook Lee, S. Rha, B. Shim, William Ho, M. Chisamore, Paul Dong Rhee, Sooa Jung, Taewan Kim, Jiyeon Yoon, Hosung Jang, E. Baek, Young-Su Noh.(2025). A phase 2 study to assess the safety and efficacy of FLX475 (tivumecirnon) combined with pembrolizumab in patients with advanced or metastatic gastric cancer..
[20] Adam Grant, Juraj Adamik, Jingtao Qiu, Jacob Wert, Molly Grandcolas, S. Jacobson, R. Goyal, William Ho, Shoji Ikeda, Marvin Au, Damian L Trujillo, M. Chisamore, D. Nagata, M. S. Ghomi, D. Brockstedt, P. Kassner, George E. Katibah.(2024). Abstract 2485: A combined mregDC and Treg signature associates with antitumor efficacy of CCR4 antagonist tivumecirnon FLX475.
[21] Y. Mahnke, K. Fletez-Brant, I. Sereti, M. Roederer.(2016). Reconstitution of Peripheral T Cells by Tissue-Derived CCR4+ Central Memory Cells Following HIV-1 Antiretroviral Therapy.
[22] J. Bogacka, Katarzyna Ciapa?a, K. Pawlik, K. Kwiatkowski, J. Dobrogowski, A. Przeklasa-Muszyńska, J. Mika.(2020). CCR4 Antagonist (C021) Administration Diminishes Hypersensitivity and Enhances the Analgesic Potency of Morphine and Buprenorphine in a Mouse Model of Neuropathic Pain.















