告別“大海撈針”:如何為研究精準鎖定最優抗標簽抗體
抗標簽抗體與表位標簽可形成嚴格的抗原-抗體特異性結合對。其中,表位標簽是經基因工程手段融合表達于目標蛋白的已知短肽,抗標簽抗體作為高特異性分子探針,能識別該短肽序列,進而完成對目標融合蛋白的定性、定量、定位及親和純化分析。在生物工程領域,二者共同搭建起標準化研究平臺。表位標簽作為融合在目標蛋白上的通用“標識”,與高特異性的抗標簽抗體協同作用,可實現對不同重組蛋白的高通量檢測與純化。該體系在生物制藥工藝監控、質量控制以及功能基因組學中蛋白互作解析等方面應用廣泛,顯著提升了研發效率與標準化程度。
1. 表位標簽
1.1 定義
表位標簽(Epitope Tag,又稱標簽肽)是一種廣泛應用于分子生物學、生物化學和細胞生物學研究中的技術工具。它指的是將一段已知的、較短的外源多肽序列(通常為8-15個氨基酸)與目標蛋白(N端或C端)人為融合表達,從而利用針對該標簽的特異性抗體進行檢測、純化或定位目標蛋白。表位標簽與融合蛋白具有顯著區別:融合蛋白是一個大的概念,指通過DNA重組技術將兩個不同的基因連接在一起表達出的蛋白質(例如:GFP熒光蛋白+目標蛋白)。而表位標簽僅僅提供一段“被抗體識別的序列”,它本身不具備催化或發光等生物活性,其主要功能是作為檢測和純化的手柄。
● 表位標簽的常見特征
- 尺寸短小:通常由6-15個氨基酸組成。這種設計對目標蛋白的空間構象干擾極小,不容易影響目標蛋白的折疊、定位或生物活性。
- 特異性極高:這些序列來源于流感病毒(HA標簽)、人原癌基因(Myc標簽)等外源生物,在絕大多數實驗用的細菌、哺乳動物細胞內不存在,因此不會產生交叉反應,背景干凈。
- 免疫原性明確:雖然短,但它能被特定的單克隆抗體精準識別,親和力強。
- 位置靈活:可放置在蛋白的任意一端(甚至中間),以適應不同的實驗需求。
1.2 常見表位標簽及其特性
● 小肽標簽
| 標簽類型 | 序列與大小 | 優勢 | 局限性 | 典型應用場景 |
|---|---|---|---|---|
| FLAG標簽 | 序列: DYKDDDDK |
1. 高特異性:短序列設計減少非特異性結合,適用于免疫共沉淀(IP)、Western Blot等實驗。 2. 抗體豐富:如M2 anti-FLAG®抗體(Sigma-Aldrich)被廣泛驗證,支持多種檢測方法。 3. 可切割性:含腸激酶(Enterokinase)切割位點,便于標簽切除以獲得天然蛋白。 |
1. 分子量較小:可能影響某些小蛋白的天然構象或功能。 2. 帶負電荷:在電泳中可能影響蛋白遷移率,需優化實驗條件。 3. 成本較高:抗體價格高于部分其他標簽抗體。 |
1. 蛋白互作研究:免疫共沉淀(IP)中捕獲目標蛋白及其互作伙伴。 2. 藥物篩選:結合流式細胞術(FCM)分析細胞表面受體表達變化。 3. 基因治療:驗證病毒載體包裝蛋白的純度與活性。 |
| 大小: 8個氨基酸(約1 kDa) | ||||
| HA標簽 | 序列: YPYDVPDYA |
1. 天然來源:源自流感病毒血凝素蛋白,免疫原性低,適合活細胞成像。 2. 多應用場景:支持Western Blot、IP、流式細胞術(FCM)及免疫熒光(IF)。 3. 高親和力抗體:如12CA5抗體(Roche)靈敏度高,可檢測低豐度蛋白。 |
1. 序列較長:可能增加蛋白融合后的空間位阻,影響功能。 2. 交叉反應風險:與宿主細胞內某些蛋白存在弱結合,需設置陰性對照。 3. 切割效率低:缺乏高效酶切位點,標簽切除較困難。 |
1. 活細胞動態追蹤:免疫熒光(IF)觀察蛋白亞細胞定位(如核質轉運)。 2. 疫苗研發:檢測病毒樣顆粒(VLP)表面抗原表達。 3. 細胞信號通路:分析受體酪氨酸激酶(RTK)的內吞與降解。 |
| 大小: 9個氨基酸(約1 kDa) | ||||
| Myc標簽 | 序列: EQKLISEEDL |
1. 經典標簽:廣泛用于蛋白表達驗證、互作研究及亞細胞定位分析。 2. 多技術兼容:支持Western Blot、IP、IF、FCM及染色質免疫沉淀(ChIP)。 3. 高靈敏度抗體:如9E10抗體(Santa Cruz)可檢測納克級蛋白。 |
1. 分子量較大:可能影響小蛋白的折疊或活性。 2. 帶電荷序列:可能干擾蛋白表面電荷分布,需評估對功能的影響。 3. 批次差異:部分抗體存在批間差異,需優化實驗條件。 |
1. 轉錄因子研究:染色質免疫沉淀(ChIP)分析DNA結合活性。 2. 蛋白穩定性:通過脈沖追蹤實驗(Pulse-Chase)監測蛋白降解速率。 3. 臨床前檢測:ELISA定量檢測重組蛋白藥物在血清中的濃度。 |
| 大小: 10個氨基酸(約1.2 kDa) | ||||
| V5標簽 | 序列: GKPIPNPLLGLDST |
1. 長序列優勢:提供更多抗原表位,增強抗體結合特異性。 2. 高表達穩定性:在哺乳動物細胞中表達穩定,適合長期研究。 3. 多檢測方法:支持Western Blot、IP、IF及ELISA,且背景信號低。 |
1. 分子量較大:可能顯著改變小蛋白的物理性質(如遷移率、溶解度)。 2. 酶切位點少:缺乏通用酶切位點,標簽切除需定制化方案。 3. 應用范圍較窄:抗體選擇少于FLAG/HA標簽,需驗證抗體兼容性。 |
1. 高通量蛋白表達:在HEK293或CHO細胞中驗證重組蛋白表達水平。 2. 病毒載體研究:檢測腺相關病毒(AAV)衣殼蛋白的組裝效率。 3. 結構生物學:通過X射線晶體學或冷凍電鏡解析蛋白三維結構(需切除標簽后)。 |
| 大小: 14個氨基酸(約1.4 kDa) |
● 親和純化標簽
| 標簽類型 | 序列 | 大小 | 優勢 | 局限性 | 典型應用場景 |
|---|---|---|---|---|---|
| 6xHis標簽 | HHHHHH(6個組氨酸殘基) | 約0.8 kDa(不含連接肽) |
1. 標簽小,對蛋白結構影響小 2. 純化條件溫和 3. 成本低,操作簡單 |
1. 金屬離子可能干擾功能 2. 非特異性結合 3. 影響結晶或相互作用研究 |
1. 結構生物學研究(如X射線晶體學、冷凍電鏡) 2. 蛋白相互作用分析(如Co-IP、Pull-down) 3. 高通量表達篩選(小標簽減少干擾) |
| GST(谷胱甘肽S-轉移酶) | 約211個氨基酸(以大腸桿菌GST為例) | 約26 kDa |
1. 促進蛋白可溶性折疊 2. 純化步驟簡單 3. 標簽可切除 |
1. 標簽較大 2. 谷胱甘肽成本高 3. 融合蛋白易降解 |
1. 真核蛋白表達(如哺乳動物細胞或酵母系統) 2. 包涵體蛋白復性(GST促進正確折疊) 3. 酶活性研究(標簽可切除后分析功能) |
| MBP(麥芽糖結合蛋白) | 約370個氨基酸 | 約42.5 kDa |
1. 顯著提高溶解性 2. 純化步驟簡單 3. 標簽可切除 |
1. 標簽大 2. 直鏈淀粉樹脂成本高 3. 洗脫條件可能影響活性 |
1. 膜蛋白或難表達蛋白(MBP顯著提高溶解性) 2. 功能研究(如酶活性、結合能力測試) 3. 疫苗開發(MBP融合抗原增強免疫原性) |
● 熒光蛋白標簽
① GFP及其變體
GFP(綠色熒光蛋白):源自水母,由238個氨基酸組成(約27 kDa),在藍光激發下發出綠色熒光(激發峰488 nm,發射峰507 nm)。
變體優化:
- EGFP(增強型GFP):通過F64L和S65T突變,熒光強度提升35倍,光穩定性增強,成熟速度加快,適用于37℃哺乳動物細胞表達。
- Emerald GFP:進一步優化光穩定性和成熟速度,適合動態過程追蹤(如細胞分裂)。
- Superfolder GFP:耐酸性強,折疊效率高,適用于易形成包涵體的融合表達。
- CFP(青色熒光蛋白):激發峰433/453 nm,發射峰475/510 nm,常作為FRET供體與YFP配對。
② YFP
YFP(黃色熒光蛋白):發射光譜紅移至黃色區域(激發峰513 nm,發射峰527 nm),用于區分GFP信號。
優勢
- 非侵入性實時成像:無需固定細胞或添加底物,直接通過熒光顯微鏡觀察活細胞內蛋白動態。
- 高靈敏度與低背景:自發熒光特性避免細胞內其他產物干擾,適用于高通量藥物篩選。
- 多色成像能力:與RFP、YFP等變體組合,實現多目標同時追蹤(如共定位分析)。
局限性
- 標簽效應:可能改變目標蛋白的相分離行為(如GFP標記的Dhh1提高相分離臨界濃度),需通過未標記對照驗證。
- 光毒性:長時間激發可能導致細胞損傷,需優化光照劑量(如使用LED光源)。
- 光譜重疊:部分變體(如YFP與GFP)光譜重疊,限制多色成像分辨率。
典型應用場景
- 蛋白定位與動態追蹤:標記微管蛋白觀察細胞分裂過程。
- 基因表達報告:將GFP置于啟動子下游,實時監測基因激活狀態。
- 蛋白質相互作用研究:通過FRET技術檢測鈣調蛋白與靶蛋白結合(如CFP-YFP組合)。
③ RFP和mCherry
RFP(紅色熒光蛋白):從珊瑚中克隆(如DsRed為225個氨基酸),發射紅色熒光(激發峰558 nm,發射峰583 nm),但成熟慢且易形成四聚體。
mCherry:DsRed的優化變體,單體形式(236個氨基酸),激發峰587 nm,發射峰610 nm,具有高亮度與光穩定性、快速成熟、低背景干擾等優點,且因單體形式可避免四聚體(如DsRed)導致的蛋白融合干擾。
優勢
- 長波長激發:減少細胞自發熒光干擾,適合深層組織成像。
- 廣泛適應性:已用于原核和真核細胞(如哺乳動物細胞、酵母)。
- 操作簡便:表達系統無需誘導劑,適用于誘導劑添加困難場景(如重組菌在實驗動物胃腸道中表達蛋白)。
- 兼容性:可與GFP或其他報告基因結合,實現多色標記和共定位研究。
局限性
- 標簽效應:雖為單體,但融合表達時仍可能輕微影響目標蛋白功能(需通過未標記對照驗證)。
- 成本:RFP相關試劑(如抗體)成本較高。
- 光譜限制:與遠紅外熒光蛋白(如iRFP)相比,組織穿透性較弱。
典型應用場景
- 長期追蹤:利用光穩定性強的特性,觀察細胞周期或基因表達動態變化。
- 多色成像:與GFP組合,區分不同蛋白或細胞結構(如內質網與線粒體共定位)。
- 流式細胞術與免疫熒光:通過抗RFP抗體檢測或擴增熒光信號,分析細胞表型或蛋白表達水平。
④ 熒光蛋白標簽在活細胞成像中的綜合應用:
多色成像策略
- 組合選擇:GFP(藍色通道)與mCherry(紅色通道)組合,避免光譜重疊,實現高分辨率共定位分析。
- 應用案例:同時標記微管蛋白(GFP)和線粒體(mCherry),觀察細胞分裂過程中線粒體分布變化。
動態過程監測
- FRAP(光漂白后熒光恢復):標記GFP融合蛋白,通過漂白區域熒光恢復速率分析蛋白轉運動力學。
- FRET技術:利用CFP-YFP組合檢測蛋白質相互作用(如鈣離子濃度變化)。
深層組織成像
- 雙光子顯微鏡:結合長波長激發的RFP(如mCherry),減少光散射,實現深層組織成像(如腦切片)。
- 近紅外熒光蛋白(iRFP):發射波長650-900 nm,組織穿透性更強,適用于體內成像(如腫瘤模型)。
超分辨成像技術
- STED顯微鏡:利用熒光蛋白的熒光壽命差異,突破衍射極限,解析凝聚體內部亞結構(如亨廷頓蛋白聚集體)。
- 單分子定位顯微術(SMLM):通過光激活或光轉換蛋白(如PA-GFP、Kaede),實現納米級分辨率成像。
值得一提的是,除了直接使用熒光蛋白,像SNAP-tag和HaloTag這類蛋白標簽技術也在不斷發展。它們能特異性結合外源性熒光染料,兼具了基因編碼的靶向性和化學染料的優良光學性能,為活細胞成像提供了更多選擇。
2. 抗標簽抗體和表位標簽的識別機制
抗標簽抗體是專門針對特定表位標簽開發的高度特異性抗體。其工作機制基于經典的抗原-抗體特異性識別原理.在標簽 - 抗體結合的識別機制方面,抗標簽抗體主要通過三維構象識別發揮作用,而非簡單讀取一級序列。表位標簽雖是線性氨基酸序列,但在目標蛋白表面會折疊成特定二級結構(如螺旋或轉角),抗體的抗原結合片段(Fab)識別的是其三維空間形狀及表面電荷分布,且該識別不依賴目標蛋白自身功能,只取決于標簽是否暴露。同時,為避免假陽性,抗標簽抗體通常具備極高的序列特異性。二者協同使得研究人員無需為目標蛋白逐一開發特異性抗體,即可利用同一套工具完成對任何帶標簽蛋白的檢測(如Western Blot)、可視化定位(如免疫熒光)和親和純化(如免疫沉淀),極大地提高了實驗的標準化程度和效率。
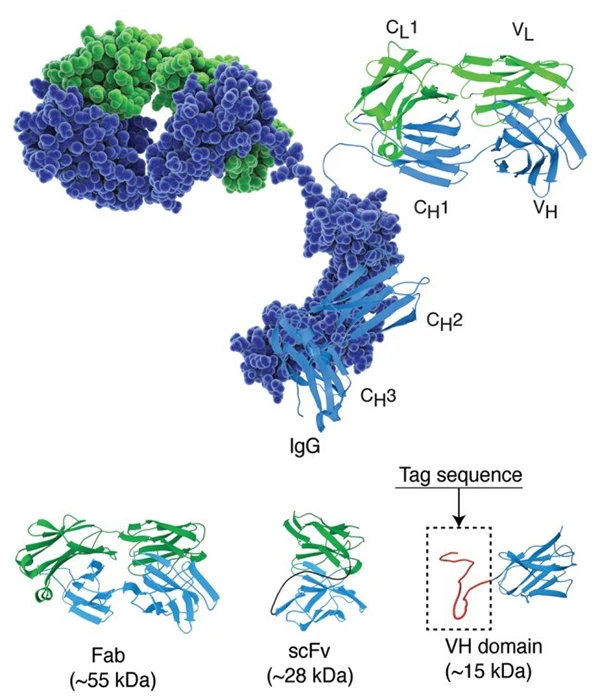
圖片來源:PMID: 25426869
譯:免疫球蛋白 G 的結構在空間填充和卡通表示中,輕鏈和重鏈分別以綠色和藍色著色。下圖說明了抗體片段 FAB(片段抗原結合)、scFv(單鏈片段可變)和單域抗體的晶體結構。大約 30 個殘基的肽標簽顯示在域抗體的 c 末端。此圖中使用的免疫球蛋白結構基于 RCSB 蛋白質數據庫條目 1igt。
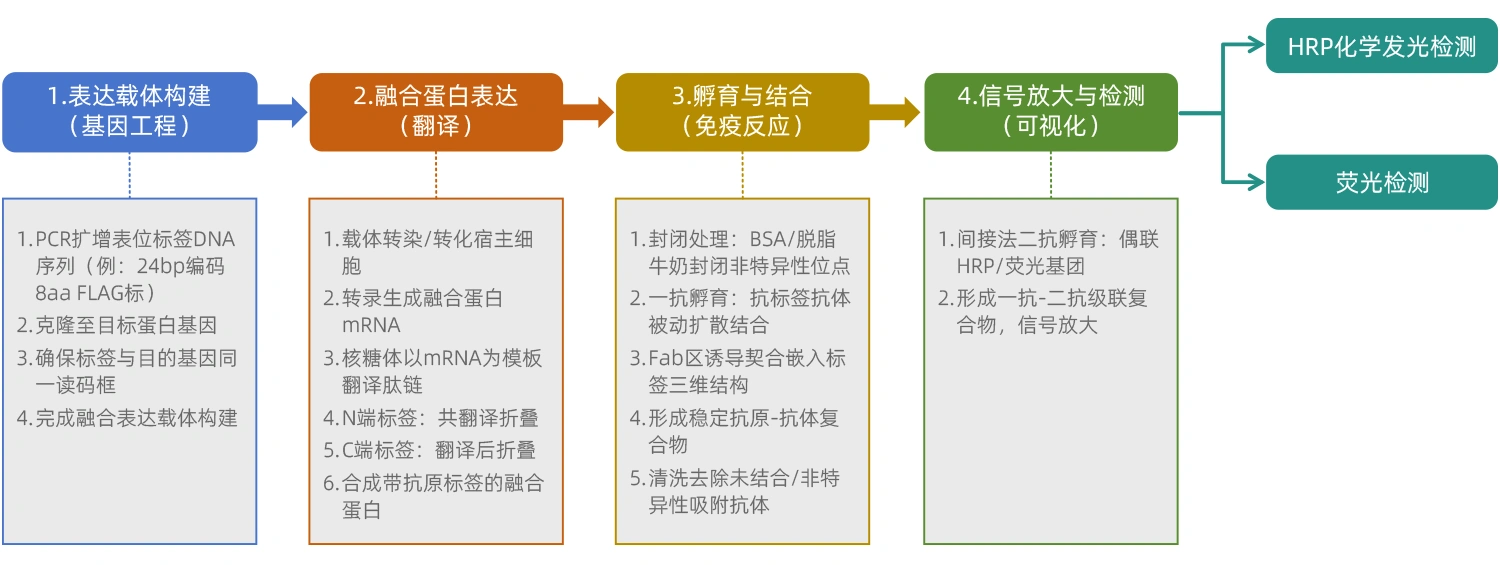
● 具體檢測流程
從標記蛋白表達到抗體識別,通常分為四個關鍵步驟:
3. 抗標簽抗體 vs 蛋白特異性抗體
| 對比維度 | 抗標簽抗體 | 蛋白特異性抗體 |
|---|---|---|
| 開發目標 | 針對標準化標簽序列(如FLAG、His)開發 | 針對特定天然蛋白質的獨特表位 |
| 開發周期與成本 | 開發周期短(2-3周),成本低,依賴雜交瘤技術或重組表達平臺,批次穩定性高。 | 開發周期長(數月),成本高,需多輪驗證特異性。 |
| 適用范圍 | 任何帶有該標簽的融合蛋白(通用工具) | 僅識別特定的天然蛋白質(專用工具) |
| 檢測原理 |
|
|
| 應用場景 | 通用檢測/純化:適用于Western Blot、免疫熒光、免疫沉淀、親和層析等,支持高通量實驗及產業化生產(如抗體藥物純化、酶制劑生產、重組蛋白表達監測、蛋白互作初篩)。 | 特異性研究:適用于蛋白互作網絡解析、信號通路研究、疾病相關蛋白檢測、天然構象分析等,需區分內源/外源性蛋白的場景。 |
| 核心優勢 |
|
|
| 局限性 |
|
|
4. 為何在研究中使用抗標簽抗體?
抗標簽抗體作為一種針對人工引入的短肽序列(即“標簽”)的特異性抗體,在生物醫學研究中已成為一種極其通用和強大的工具。其核心優勢在于,研究者無需為每一個目標蛋白從頭開發和驗證特異性抗體,而是可以利用已有的、經過高度優化的抗標簽抗體系統,實現對多種不同蛋白的檢測、純化、定位和功能操控。以下將詳細闡述其使用場景與核心優勢。
4.1 抗標簽抗體主要使用場景
● 蛋白質結晶與結構解析
在結構生物學中,獲得高質量的蛋白質晶體是重大挑戰。抗標簽抗體可以作為“結晶伴侶”來促進目標蛋白的結晶。例如,識別PA標簽的單克隆抗體NZ-1的Fab片段,可以與插入PA標簽的目標蛋白形成復合物,從而促進共結晶[1]。這種方法繞過了為每個目標蛋白開發構象特異性抗體的困難,通過將標簽插入目標蛋白的特定環區(如β-發夾結構),利用抗體與標簽的高親和力結合,穩定蛋白構象,最終獲得可用于高分辨率結構解析的晶體。
● 蛋白質檢測與可視化
- 免疫印跡(Western Blot)與免疫熒光:用于檢測和定量外源表達的標簽蛋白。
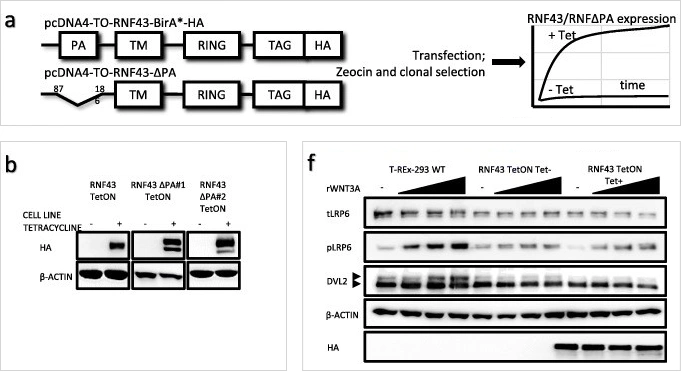
圖片來源:PMID: 30666745
圖片說明:T-REx 293 細胞系中的誘導型 RNF43 和 RNF43ΔPA 表達。 a.所用實驗模型的示意圖。b.Western 印跡顯示 Tet 誘導的 HA 標記的 RNF43 構建體在穩定細胞系中的表達。f. 通過應用濃度增加的經典 Wnt 通路激活的蛋白質印跡分析處理
- 免疫電子顯微鏡:用于在超微結構水平上定位膜蛋白復合物。通過工程化的抗體片段(Fv),其一端結合目標蛋白的表位,另一端攜帶標簽(如Strep標簽或c-myc標簽),再通過對應的檢測系統(如膠體金標記的鏈霉親和素或抗標簽抗體)實現高分辨率定位。
- 免疫組織化學/細胞化學:基本原理同上,用于在組織或細胞切片中定位標簽蛋白。
● 染色質免疫共沉淀
染色質免疫共沉淀是研究蛋白質與DNA體內相互作用的關鍵技術。當針對內源轉錄因子的特異性抗體不可得或效果不佳時,表達外源的表位標簽融合蛋白,并使用高特異性的抗標簽抗體進行免疫沉淀,成為一種可靠且高效的替代方案[2]。這種方法避免了耗時費力地制備轉基因穩定株系,例如在擬南芥原生質體中瞬時表達標簽蛋白即可在4天內完成ChIP實驗。
● 蛋白質純化
抗標簽抗體可用于快速、一步法親和純化重組蛋白。例如,Sanae Tabata等人于2010年利用抗TARGET標簽抗體P20.1,可以建立靈敏的篩選體系,從哺乳動物細胞培養上清中快速篩選高產細胞系,并標準化地純化出毫克級的靶蛋白。同樣,在Fv片段的生產中,通過其融合的Strep標簽也可進行一步親和純化。
● 功能研究與調控
抗標簽抗體不僅能“看”和“抓”,還能“干擾”和“激活”蛋白功能。
- 功能干擾:某些抗標簽抗體的結合可以阻斷蛋白質相互作用。例如,一種抗SV40大T抗原的抗體,因其表位與T抗原結合RPA70N的位點重疊,能夠直接阻斷二者的相互作用。
- 功能激活:在合成生物學應用中,抗標簽抗體可用于人工誘導蛋白質二聚化以激活信號。例如,在原型囊泡內表達的跨膜融合蛋白,其胞外端帶有標簽;當外部添加對應的抗標簽抗體時,可引起胞內報告酶(GUS)的條件性二聚化與激活,從而實現信號放大與檢測。
● 診斷與檢測技術開發
抗標簽抗體系統為高靈敏度檢測平臺的構建提供了模塊化工具。
- 數字ELISA與生物傳感器:基于原型囊泡的陣列利用抗標簽抗體誘導的跨膜信號傳導,實現了對特定抗體(如曲妥珠單抗)甚至小分子抗原(如咖啡因)的高靈敏度、數字化檢測,且減少了繁瑣的洗滌步驟。
- 磁弛豫開關:在基于磁性顆粒的傳感中,通過使用抗標簽抗體及其二抗來增加靶標效價,可以顯著增強檢測靈敏度。
- 蛋白芯片:在自組裝蛋白芯片技術中,首先將抗標簽抗體打印在芯片上,然后利用無細胞表達系統在芯片原位合成標簽融合蛋白,并立即被抗體捕獲,從而高通量地生產功能蛋白微陣列。
● 臨床診療應用
在臨床領域,放射性核素標記的抗標簽抗體可用于腫瘤的定位與手術導航。例如,在復發性結直腸癌手術中,使用碘-125標記的抗TAG抗體(CC49單克隆抗體)進行放射免疫引導手術,能夠發現更多傳統影像學和手術探查無法發現的隱匿病灶,從而改變手術方案,提高腫瘤切除率。
4.2 核心優勢
綜合以上場景,抗標簽抗體的優勢主要體現在以下幾個方面:
- 通用性與高效性:一套成熟的抗標簽抗體系統(如抗HA、myc、FLAG、PA標簽的抗體)可以應用于任何被相應標簽標記的蛋白,無需為每個新蛋白定制抗體,極大地節省了時間與成本。
- 高特異性與高親和力:許多商業化的抗標簽抗體經過精心篩選和優化,具有極高的特異性和親和力。
- 靈活性:標簽可以相對自由地插入目標蛋白的N端、C端或內部環狀區域,以適應不同的實驗需求,如表面展示、避免干擾功能域等。
- 提高實驗成功率與可重復性:使用經過驗證的抗標簽抗體進行ChIP、純化或檢測,可以避免因內源抗體質量不佳而導致的實驗失敗,結果更加穩定可靠。
- 實現創新性實驗設計:抗標簽抗體為前沿技術提供了關鍵組件,例如在無細胞表達系統中構建蛋白芯片、在合成囊泡中構建信號轉導系統,以及開發新型生物傳感器等,推動了方法學的進步。
5. 新興標簽技術
5.1 新一代小標簽HiBiT及其他超小標簽
傳統蛋白質標簽(如GFP、HaloTag)雖然功能強大,但其較大的分子量(通常超過20 kDa)可能干擾目標蛋白(POI)的天然結構、功能、定位和相互作用。新一代超小標簽技術,以HiBiT肽為代表,正在解決這些痛點。
HiBiT標簽的核心優勢在于其極小的尺寸和高亮度的互補發光系統。 HiBiT是一個僅由11個氨基酸組成的短肽(約1.3 kDa),它本身不發光,但能與一個較大的互補蛋白片段(LgBiT,18 kDa)以極高的親和力結合,形成完整的、具有活性的NanoLuc熒光素酶。這一設計帶來了多重革命性優勢:
- 最小化對目標蛋白的干擾:由于HiBiT標簽極小,將其融合到目標蛋白的N端或C端,對蛋白的天然折疊、膜定位、信號轉導及蛋白-蛋白相互作用的干擾遠小于傳統大標簽。這使得在近乎天然的生理環境下研究蛋白質成為可能,數據更具生理相關性 [3]。
- 實現高信噪比的細胞表面選擇性檢測:LgBiT蛋白無法穿透細胞膜。因此,只有當HiBiT標簽位于活細胞表面時,才能與外加的LgBiT互補產生發光信號。這一特性被巧妙用于實時、動態監測細胞表面受體的數量變化,例如G蛋白偶聯受體(GPCR)在配體刺激后的內吞過程。該方法背景極低,因為細胞內的受體無法產生信號 [4]。
- 超高靈敏度和寬動態范圍:NanoLuc熒光素酶是已知最亮的熒光素酶之一。HiBiT/LgBiT互補系統繼承了這一高亮度特性,能夠檢測到極低豐度的目標蛋白,甚至適用于表達量極低的外源蛋白研究模型。其寬動態范圍便于精確量化從微弱到強烈的信號變化 [4]。
- 支持多重檢測與動態過程分析:HiBiT系統兼容其他標記技術(如HaloTag、SNAP-tag),為多色單分子成像和同時追蹤多種蛋白質的動態提供了可能2。此外,基于HiBiT的分泌報告系統能夠以高靈敏度、高通量的方式實時監測細菌III型分泌系統等動態生物學過程 [5]。
- 簡化病毒學與藥物篩選研究:HiBiT標簽可被插入到病毒蛋白(如肝炎E病毒ORF2蛋白)的特定位點,構建出帶有發光報告基因的感染性病毒8。這種重組病毒允許方便、定量地實時監測病毒感染和復制過程,極大地促進了抗病毒藥物和中和抗體的評估 [6]。
5.2 納米抗體可識別的標簽
納米抗體-標簽系統的工作原理是駱駝科動物或人工設計單域抗體,能高親和力、特異性結合6 - 14個氨基酸短肽序列,短肽標簽可融合到目標蛋白N端或C端,表達后納米抗體可作通用工具用于蛋白捕獲、可視化或操縱。
● 納米抗體可識別標簽類型
- 高特異性短肽標簽:①.6 - 7個氨基酸短肽標簽:基于納米抗體的高效純化系統,能一步純化多種可溶性和膜蛋白,純度與回收率高,結合蛋白可通過競爭性合成肽溫和洗脫,回收率超90%。②.14 - mer肽表位標簽(如Nb6E系統):納米抗體Nb6E與14個氨基酸肽表位相互作用,鑒定出關鍵殘基并提出相互作用模式,用于增強工程化腺苷A2A受體配體效力 [7]。
- 用于共價交聯的標簽系統:合成肽表位與納米抗體配對,在活細胞表面實現快速、特異共價交聯,交聯反應速率快,可用于標記活細胞并研究配體交聯對受體信號傳導的影響
- 用于蛋白質定位與鄰近標記的標簽系統:ALFA標簽系統,基于ALFA標簽 - 納米抗體配對,用于分枝桿菌中蛋白質定位和鄰近蛋白質組學研究,適用于多種微生物系統
- 用于分泌生產的C端識別序列: marcescens的LipC分泌系統識別納米抗體C末端Val - Thr - Val序列,將納米抗體單步分泌至胞外,構成基于特定序列的分泌識別標簽
● 納米抗體-標簽系統的優勢
- 高親和力與特異性:亞納摩爾甚至皮摩爾級別結合,背景干擾低
- 標簽極小:對目標蛋白結構和功能影響小
- 溫和洗脫:利于保持目標蛋白活性
5.3 TEV蛋白酶切割位點
盡管小標簽和納米抗體的干擾已降至最低,但在某些精細研究中,完全移除標簽以獲得與天然狀態完全一致的蛋白質產物仍然是必要的。可切割標簽系統應運而生,其中,TEV蛋白酶切割位點是最常用和高效的系統之一。
TEV蛋白酶是一種高度特異性的蛋白酶,能識別其特定的七氨基酸序列(Glu-Asn-Leu-Tyr-Phe-Gln-Gly或類似變體),并在Gln與Gly/Ser之間進行切割。 在蛋白純化或功能研究流程中,何時需要引入并最終移除標簽主要基于以下考量:
- 獲得無標簽的純化蛋白:在重組蛋白表達中,常使用較大的親和標簽(如His-tag、GST-tag)來簡化純化過程。在親和標簽與目標蛋白之間引入TEV蛋白酶位點,便可在純化后通過酶切獲得高純度的無標簽目標蛋白。
- 控制蛋白質的激活或釋放:在合成生物學或細胞工程中,可切割標簽可用于設計條件性激活的蛋白質。例如,將某個功能域或抑制肽通過TEV位點與主蛋白連接,使其處于失活狀態。只有在特定條件下(如表達或加入TEV蛋白酶),切割發生,功能域被釋放,蛋白質才被激活。
- 解決研究模型中的特殊需求:在標簽抗體與目標病毒蛋白之間設計可切割連接子,可在完成報告基因檢測的初步研究后,通過切割獲得不含報告基因的病毒蛋白,用于更深入的功能驗證。
6. 如何選擇合適的抗標簽抗體
在生物工程實驗中,抗標簽抗體是連接目標蛋白與檢測技術的關鍵工具。然而,面對市場上琳瑯滿目的標簽抗體產品,如何根據實驗需求精準選擇?我們可以從實驗目標、標簽設計、抗體特性到技術兼容性,系統梳理抗標簽抗體的選擇策略,助您提升實驗效率與數據可靠性。
6.1 基于實驗目標選擇標簽抗體
● 蛋白質定位研究:熒光或小肽標簽是首選
蛋白質定位研究需通過熒光信號或免疫染色直觀呈現目標蛋白的亞細胞分布。此時,熒光蛋白標簽(如GFP、mCherry)或小肽標簽(如HA、FLAG)更為適用:
熒光標簽:直接提供可視化信號,無需額外染色步驟,適合活細胞成像,需匹配顯微鏡激發光源(如GFP需488nm激光)。
小肽標簽:通過免疫熒光或免疫組化檢測,兼容固定細胞樣本,且標簽體積小,對蛋白功能干擾低。
● 蛋白質純化
蛋白質純化需通過標簽與配基的高親和力結合實現高效捕獲,再通過洗脫條件(如pH、競爭劑)釋放目標蛋白。需結合目標蛋白特性選擇標簽(如膜蛋白優先His標簽,易聚集蛋白優先MBP標簽),且純化后需通過酶切或化學裂解去除標簽,避免影響下游功能實驗。
常用標簽包括His、GST、MBP標簽。
His標簽:適用于金屬螯合層析(IMAC),洗脫條件溫和(如咪唑梯度),但需注意宿主蛋白內源性組氨酸的干擾。
GST標簽:通過谷胱甘肽洗脫,適合大規模純化,但標簽較大(26 kDa)可能影響蛋白溶解性。
MBP標簽:增強目標蛋白可溶性,通過麥芽糖洗脫,適用于難折疊蛋白。
● 蛋白質-蛋白質相互作用研究
相互作用研究需通過免疫共沉淀(Co-IP)或Pull-down技術捕獲蛋白復合物,標簽需滿足:
小體積:避免空間位阻干擾天然相互作用(如FLAG、HA、Myc標簽僅含8-10個氨基酸)。
高特異性:抗體需嚴格識別標簽序列,避免交叉反應。
FLAG、HA、Myc標簽的可獲得最小化干擾。
6.2 標簽大小與位置
● 標簽大小對蛋白質功能的影響
標簽分子量建議不超過目標蛋白的10%(如50 kDa蛋白優先選擇<5 kDa的標簽)。標簽體積過大可能干擾目標蛋白的折疊、穩定性或相互作用界面。實操過程中可通過AlphaFold預測標簽與目標蛋白的接觸面,避免覆蓋關鍵功能域。
● N端 vs. C端標記
N端標記:適用于無信號肽的胞質蛋白,但可能干擾分泌蛋白的轉運(如抗體輕鏈N端標記會阻斷分泌)。
C端標記:更通用,但需避免破壞蛋白質C端的定位信號(如線粒體靶向序列)。
內部標記:對長蛋白可在柔性 loop區插入標簽,需通過突變庫篩選不影響功能的位點,內部標記適用于目標蛋白N/C端功能關鍵(如酶活性中心)。
6.3 抗體特性:特異性、靈敏度與驗證數據
● 單克隆 vs. 多克隆抗體
單克隆抗體:特異性強、批次差異小,適合定量實驗(如WB、ELISA),但開發成本高。
多克隆抗體:靈敏度高、可識別多個表位,適合低豐度蛋白檢測,但批次間差異需嚴格質控。
關鍵實驗(如臨床樣本檢測)優先單克隆抗體。預實驗或篩查階段可使用多克隆抗體降低成本。
● 靈敏度與特異性
靈敏度:根據目標蛋白表達量選擇抗體(如低表達蛋白需高親和力抗體,如KD<1nM)。
特異性:通過WB檢測目標蛋白與宿主蛋白(如E.coli、HEK293)的交叉反應,優先選擇經敲除細胞系驗證的抗體。
● 應用驗證
驗證數據:檢查抗體是否通過WB、IP、IF、ChIP等多技術驗證,避免使用僅通過ELISA驗證的抗體用于IP實驗。
● 宿主物種選擇
一抗宿主:常用兔、鼠、羊,需與二抗宿主物種匹配(如兔抗FLAG一抗需搭配抗兔IgG二抗)。選擇不同宿主來源的一抗(如鼠抗HA+兔抗Myc),避免二抗交叉結合。
6.4 技術兼容性
● 固定方法兼容性:甲醛 vs. 甲醇
甲醛固定:交聯蛋白,可能掩蓋線性表位,需選擇識別構象表位的抗體(如某些熒光標簽抗體)。
甲醇固定:適用于膜蛋白,但可能破壞某些標簽的抗原性(如GST標簽在甲醇中易變性)。
● 抗原修復要求:熱誘導 vs. 酶法
熱修復:適用于甲醛固定的石蠟切片,通過高溫恢復表位(如100℃加熱20分鐘)。
酶修復:使用蛋白酶K或胃蛋白酶,需優化濃度與時間以避免過度消化。
● 天然 vs. 變性蛋白識別(構象表位 vs. 線性表位)
天然蛋白:需選擇識別構象表位的抗體(如某些熒光標簽抗體僅在蛋白正確折疊時結合)。
變性蛋白:適用于WB實驗,需選擇識別線性表位的抗體(如His標簽抗體在SDS-PAGE中仍可結合)。
6.5 串聯標簽策略
● 多個相同標簽的協同效應
例如在目標蛋白兩端各插入一個FLAG標簽(FLAG-蛋白-FLAG),通過雙抗體夾心法提升檢測信號。His6標簽串聯(His6-His6)可提高與鎳柱的結合容量,適合低表達蛋白純化。
● 不同標簽的組合應用
純化+檢測標簽:如His標簽用于純化,V5標簽用于WB檢測,避免純化標簽干擾下游分析。
多色實驗標簽:如GFP(定位)+HA(相互作用)+Myc(表達量),實現多維度信息獲取。
7. 實驗設計最佳實踐
7.1 設計標記融合蛋白
● 標簽插入位置原則
目標:平衡蛋白質功能、抗體可及性與結構完整性。
操作步驟:
- 功能域分析:使用UniProt、PDB數據庫或AlphaFold預測目標蛋白的結構域與信號序列。
- 抗體結合可及性:優先選擇蛋白質表面暴露的柔性區域(如無規則卷曲區)插入標簽。通過PyMOL或ChimeraX可視化蛋白結構,標記潛在插入位點。
- 信號序列規避:分泌蛋白(如抗體)需保留N端信號肽,標簽應插入信號肽下游(如IgG輕鏈N端信號肽后接His標簽)。
● 使用接頭序列
目標:減少標簽對目標蛋白構象的干擾,提升抗體結合效率。
操作步驟:
- 柔性接頭添加條件:標簽與目標蛋白直接連接可能導致空間位阻(如FLAG標簽與膜蛋白跨膜區相鄰時)。
- 接頭長度與組成優化:
長度:通常4-20個氨基酸,短接頭(如GGGGS)適用于小標簽(如His),長接頭(如(GGGGS)?)適用于大標簽(如GST)。組成:避免富含脯氨酸(Pro)或帶電氨基酸(如Arg、Lys),推薦使用甘氨酸(Gly)和絲氨酸(Ser)柔性組合。
● 表達系統考慮
目標:根據蛋白復雜度選擇表達系統,平衡產量與功能正確性。
操作步驟:
- 細菌 vs. 哺乳動物表達:
細菌表達(如E. coli):適合無翻譯后修飾的簡單蛋白(如His-ERK2),成本低但易形成包涵體。哺乳動物表達(如HEK293):適用于需糖基化或二硫鍵的蛋白(如膜受體-GFP融合),但成本較高。
- 標簽切除選項:
酶切位點設計:在標簽與目標蛋白間插入TEV或Thrombin識別序列(如ENLYFQG或LVPRGS),純化后通過酶切去除標簽。自切割標簽:使用intein標簽(如IMPACT系統),通過pH或溫度誘導自動切除。
7.2 設置適當的對照
● 陰性對照
目標:排除非特異性背景信號。
操作步驟:
- 未轉染細胞:檢測細胞本底熒光或抗體非特異性結合(如流式細胞術中未轉染細胞的自發熒光)。
- 空載體對照:轉染不含目標蛋白的載體(如pcDNA3.1空載體),驗證標簽抗體無交叉反應。
● 同型對照
目標:校正抗體非特異性結合(尤其在IF和流式中)。
操作步驟:
- 同型抗體選擇:與一抗同種屬、同亞型但無關靶標的抗體(如抗FLAG一抗為兔IgG,同型對照用兔IgG同型匹配抗體)。
- 濃度匹配:同型對照濃度與一抗相同(如1 μg/mL)。
● 陽性對照
目標:驗證實驗體系有效性。
操作步驟:
- 已知標記蛋白:使用已驗證的標記蛋白(如商業化GFP-actin融合蛋白)作為WB或IF陽性對照。
- 商業化對照裂解物:使用含目標蛋白的細胞裂解物(如HeLa細胞裂解物檢測內源性ERK2)。
8. 常見問題故障排除
| 故障類型 | 可能原因 | 現象分析 | 解決方法 |
|---|---|---|---|
| 無信號或信號弱 | 標記蛋白表達量低 | 內參條帶正常,目的蛋白無信號。 | 驗證轉染/轉導效率:使用熒光共轉染或流式細胞術確認陽性細胞比例。 |
| 優化表達條件:進行誘導時間/濃度梯度實驗,更換啟動子。 | |||
| 檢查蛋白質穩定性:使用蛋白酶體抑制劑(如MG132)處理細胞,觀察信號是否恢復。 | |||
| 抗體濃度不足 | 陽性對照有微弱信號,樣品無信號。 | 滴定實驗:對同一樣品膜使用不同稀釋度(如1:500, 1:1000, 1:5000)孵育,尋找最佳濃度。 | |
| 推薦起始稀釋度:新抗體從說明書范圍中間值開始(如1:100-1000則取1:500)。 | |||
| 標簽表位被屏蔽 | 標簽抗體無信號,但目的蛋白特異性抗體有信號。 | 蛋白質折疊:提高樣品煮沸溫度(95°C,10 min)或增強變性劑強度,使表位充分暴露。 | |
| 替代標簽位置:若N端標簽被屏蔽,嘗試構建C端標簽的質粒。 | |||
| 轉印效率問題 | 凝膠染色有殘留蛋白,或膜上Marker不清。 | 蛋白質大小考慮:大分子蛋白(>150 kDa)選用濕轉、低甲醇、長時間;小分子蛋白(<20 kDa)選用0.2μm孔徑膜、短時間。 | |
| 轉印優化:轉印后常規進行麗春紅染色,驗證轉印效果并觀察上樣一致性。 | |||
| 高背景信號 | 抗體濃度過高 | 整張膜模糊一片,無清晰條帶。 | 優化策略:降低一抗或二抗濃度,縮短孵育時間。特別注意二抗濃度通常是高背景的主要來源。 |
| 封閉不充分 | 背景均勻,呈磨砂狀。 | 封閉緩沖液選擇:常規用5%脫脂牛奶;檢測磷酸化蛋白時改用5% BSA(避免牛奶中酪蛋白干擾)。 | |
| 封閉時間和溫度:室溫至少1小時,或4°C過夜。 | |||
| 洗滌不充分 | 背景呈局部塊狀或條紋狀污漬。 | 洗滌緩沖液成分:常規使用TBST(含吐溫20)。 | |
| 洗滌時長和頻率:強調“少量多次”,每次5-10分鐘,重復5次以上。 | |||
| 非特異性結合 | 除了目標條帶,還有明顯的非預期條帶背景。 | 抗體質量問題:選用親和純化抗體或文獻驗證過的單克隆抗體。 | |
| 緩沖液優化:提高抗體稀釋液/洗滌液中的鹽濃度或吐溫20含量,減弱疏水相互作用。 | |||
| 檢測到多條帶 | 蛋白質降解 | 出現比目標蛋白小的條帶(呈階梯狀)。 | 蛋白酶抑制劑的使用:裂解液必須新鮮加入蛋白酶抑制劑混合物(Cocktail)。 |
| 樣品處理方案:全程冰上操作,裂解后立即煮沸并快速冷凍,避免反復凍融。 | |||
| 翻譯后修飾 | 出現比目標蛋白大的條帶,或有規律的多條帶。 | 預期的修飾:查閱文獻確認已知修飾。進行去修飾處理(如去磷酸化酶)作為對照驗證。 | |
| 遷移率偏移:磷酸化/糖基化等修飾會改變蛋白遷移速率,屬正常現象。 | |||
| 非特異性結合 | 條帶很多,連內參通道都有雜帶。 | 抗體濃度調整:降低抗體濃度,進行滴定實驗。 | |
| 額外對照:設置敲低(Knockdown)或敲除(Knockout)樣品對照。若雜帶在敲除樣品中仍存在,則為非特異性條帶。 | |||
| 標簽特定問題 | HA標簽在凋亡細胞中被切割 | 除全長蛋白外,多出一條清晰的小片段。 | 半胱天冬酶切割位點:HA序列包含Caspase識別位點,凋亡時被切割。 |
| 預防策略:進行凋亡實驗時,改用Flag或Myc標簽;或同時在C端加標簽驗證全長蛋白。 | |||
| Myc標簽識別可變性 | Western Blot可檢測,但免疫熒光(IF)或免疫沉淀(IP)檢測不到。 | 固定依賴性表位可及性:Myc表位在特定固定液(如甲醇)或天然構象下被掩蓋。 | |
| 解決方法:IF實驗嘗試更換透膜劑(如Triton X-100)或固定液(如4% PFA)。 | |||
| His標簽非特異性結合 | 檢測或純化時背景很高,出現很多雜蛋白。 | 金屬離子污染:宿主自身含組氨酸簇的蛋白會結合鎳柱。 | |
| 緩沖液成分調整:在結合/洗滌緩沖液中添加低濃度咪唑(20-40 mM)競爭洗脫雜蛋白。 |
9. 常見問題解答(FAQ)
參考文獻
[1] Tamura R, Miyazaki N, Ito Y, et al. Application of the NZ-1 Fab as a crystallization chaperone for PA tag-inserted target proteins. Protein Sci 2019; 28: 1450–59.
[2] Fang W, Wang X, Li Y, et al. Optimized protocols for chromatin immunoprecipitation of exogenously expressed epitope-tagged proteins. STAR Protoc 2023; 4: 102050.
[3] Hanl M, Li J, Schmidt T, et al. Target Engagement Studies and Kinetic Live-Cell Degradation Assays Enable the Systematic Characterization of Histone Deacetylase 6 Degraders. ACS Pharmacol Transl Sci 2025; 8: 2456–68.
[4] Boursier ME, Levin S, Zimmerman K, et al. The luminescent HiBiT peptide enables selective quantitation of G protein-coupled receptor ligand engagement and internalization in living cells. J Biol Chem 2020; 295: 5124–35.
[5] Yoda T, Shirai Y, Okada Y, et al. Four-color single-molecule imaging system for tracking GPCR dynamics with fluorescent HiBiT peptide. Biophys Physicobiol 2024; 21: e210020.
[6] Ankavay M, Dubuisson J, Cocquerel L, et al. Monitoring of hepatitis E virus infection and replication by functional tagging of the ORF2 protein. JHEP Rep 2025; 7: 101293.
[7] Cabalteja CC, Mihaylov D, Cubitt B, et al. Characterization of a Nanobody-Epitope Tag Interaction and Its Application for Receptor Engineering. ACS Chem Biol 2022; 17: 2345–56.











