Recombinant Mouse Transcription factor Sp1 (Sp1)
-
中文名稱:小鼠Sp1重組蛋白
-
貨號:CSB-YP022438MO
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:小鼠Sp1重組蛋白
-
貨號:CSB-EP022438MO
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:小鼠Sp1重組蛋白
-
貨號:CSB-EP022438MO-B
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:小鼠Sp1重組蛋白
-
貨號:CSB-BP022438MO
-
規格:
-
來源:Baculovirus
-
其他:
-
中文名稱:小鼠Sp1重組蛋白
-
貨號:CSB-MP022438MO
-
規格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Sp1; Transcription factor Sp1; Specificity protein 1
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
表達區域:2-784
-
氨基酸序列SDQDHSMDE VTAVVKIEKD VGGNNGGSGN GGGAAFSQTR SSSTGSSSSS GGGGGQESQP SPLALLAATC SRIESPNENS NNSQGPSQSG GTGELDLTAT QLSQGANGWQ IISSSSGATP TSKEQSGNST NGSNGSESSK NRTVSGGQYV VAATPNLQNQ QVLTGLPGVM PNIQYQVIPQ FQTVDGQQLQ FAATGAQVQQ DGSGQIQIIP GANQQIIPNR GSGGNIIAAM PNLLQQAVPL QGLANNVLSG QTQYVTNVPV ALNGNITLLP VNSVSAATLT PSSQAGTISS SGSQESSSQP VTSGTAISSA SLVSSQASSS SFFTNANSYS TTTTTSNMGI MNFTSSGSSG TSSQGQTPQR VGGLQGSDSL NIQQNQTSGG SLQGSQQKEG EQSQQTQQQQ ILIQPQLVQG GQALQALQAA PLSGQTFTTQ AISQETLQNL QLQAVQNSGP IIIRTPTVGP NGQVSWQTLQ LQNLQVQNPQ AQTITLAPMQ GVSLGQTSSS NTTLTPIASA ASIPAGTVTV NAAQLSSMPG LQTINLSALG TSGIQVHQLP GLPLAIANTP GDHGTQLGLH GSGGDGIHDE TAGGEGENSS DLQPQAGRRT RREACTCPYC KDSEGRASGD PGKKKQHICH IQGCGKVYGK TSHLRAHLRW HTGERPFMCN WSYCGKRFTR SDELQRHKRT HTGEKKFACP ECPKRFMRSD HLSKHIKTHQ NKKGGPGVAL SVGTLPLDSG AGSEGTATPS ALITTNMVAM EAICPEGIAR LANSGINVMQ VTELQSINIS GNGF
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Transcription factor that can activate or repress transcription in response to physiological and pathological stimuli. Binds with high affinity to GC-rich motifs and regulates the expression of a large number of genes involved in a variety of processes such as cell growth, apoptosis, differentiation and immune responses. Highly regulated by post-translational modifications (phosphorylations, sumoylation, proteolytic cleavage, glycosylation and acetylation). Binds also the PDGFR-alpha G-box promoter. May have a role in modulating the cellular response to DNA damage. Implicated in chromatin remodeling. Plays a role in the recruitment of SMARCA4/BRG1 on the c-FOS promoter Plays an essential role in the regulation of FE65 gene expression. Positively regulates the transcription of the core clock component ARNTL/BMAL1. Plays a role in protecting cells against oxidative stress following brain injury by regulating the expression of RNF112.
-
基因功能參考文獻:
- Sp-1 negatively regulates the expression of miR-20b in macrophages. PMID: 29263044
- an immunoprecipitation assay indicated that hypoxia triggered activation of the binding activity of Sp1 to the promoters of PARP-1 and caspase-3, which is abrogated by miR-7a/b. In summary, these findings identified miR-7a/b as protectors of cardiac remodeling and hypoxia-induced injury in H9c2 cardiomyoblasts involving Sp1 and PARP-1. PMID: 27384152
- Immunoblotting, qPCR, ChIP and siRNA-mediated gene knockdown studies revealed that the activation of phosphatidylinositol 3-kinase/protein kinase C zeta pathways in poly(I:C)-stimulated cells underlies Sp1 phosphorylation and recruitment to the mCRAMP promoter, leading to enhanced transcription PMID: 28343946
- The Genomic Context and Corecruitment of SP1 Affect ERRalpha Coactivation by PGC-1alpha in Muscle Cells PMID: 27182621
- These results suggest that Sp1 regulates gene expression of AACS in Neuro-2a cells and ketone body utilization affects the balance of histone acetylation. PMID: 29137983
- the results obtained in this study show that Znf179 autoregulation through Sp1-dependent mechanism plays an important role in neuroprotection, and NGF-induced Sp1 signaling may help attenuate more extensive (ROS-induced) damage following brain injury PMID: 27918959
- Using MA, we demonstrated Sp1 as a critical stemness-related transcriptional factor protecting GBM cells against stress- and TMZ-induced death. Thus, Sp1 inhibition may prevent recurrence of malignant cells persisting after primary therapy. PMID: 28939040
- Results suggest that 25-hydroxycholesterol (25-HC) induced the interaction between NFATc1 and Sp1, reducing the level of free Sp1 to inhibit microRNA miR-139-5p expression and promote osteoclastogenesis. PMID: 28257846
- data indicated that (-)-Epicatechin inhibited AngII-induced cardiac hypertrophy by activating the SP1/SIRT1 signaling pathway. PMID: 28420000
- This resulted in Sp1 displacement from the promoter and binding of Sp3 to it, leading to p300 recruitment and histone acetylation, activating transcription. This is the first study addressing the mechanisms of physiological TLR5 expression in the intestine. Additionally, a novel insight is gained into Sp1/Sp3-mediated gene regulation that may apply to other genes PMID: 27060138
- Dp71 expression in hepatic cells is carried out, in part, by YY1-, Sp1- and Sp3-mediated transcription from the Dp71 promoter. PMID: 27143785
- SP1 expression was up-regulated in the tubular epithelial cells after acute kidney injury induced by ischemia-reperfusion.MiR-204 regulates epithelial-mesenchymal transition by targeting SP1 in the tubular epithelial cells. PMID: 27959449
- miR-124, -128, and -137 act synergistically to regulate Sp1 expression. PMID: 26369286
- YY1 and SP1 independently and cooperatively govern the Mesp1 expression during embryogenesis. PMID: 26384464
- Taken together, these results indicate that the transcription factor Sp1 upregulates the proximal promoter activity of the mouse Col11a1 gene in chondrocytes. PMID: 26487428
- In the initial stage of myocyte differentiation, transcription of the YB-1 gene was regulated by E2F1 and Sp1, and was then gradually replaced under the control of both MyoD and myogenin. PMID: 26279143
- Data indicate that Sp1 and AP-1-related factors are involved in the regulation of MFG-E8 gene transcription by targeting their binding sites in the 5'-flanking region under physiological and inflammatory states. PMID: 25711369
- Our results unveil strikingly different recruitment mechanisms of Sp1/Sp2/Sp3 transcription factor members uncovering an unexpected layer of complexity in their binding to chromatin in vivo. PMID: 25793500
- age-dependent alteration in the Fmr-1 gene expression is associated with Sp1 interaction with Fmr-1 promoter which in turn might be related with cognitive development during brain maturation and aging. PMID: 25015265
- The results of this study suggest that SP4 and SP1 upregulation may be part of the mechanisms deregulated downstream of glutamate signalling pathways in schizophrenia PMID: 25175639
- The transcription factor SP1 is induced in brain by ischemia/reperfusion. PMID: 25445985
- Data suggest that retinoic acid and GM-CSF-induced retinal dehydrogenase 2 (RALDH2) expression in dendritic cells requires cooperative binding of transcription factor Sp1 via the RA receptor/retinoid X receptor complex to the Aldh1a2 promoter. PMID: 24788806
- Sp1/Sp3 transcription factors have roles in regulating hallmarks of megakaryocyte maturation and platelet formation and function PMID: 25538045
- Our findings suggest that an SOHLH2/SOHLH1/SP1 ternary complex autonomously and cooperatively regulates Sohlh1 gene transcription through juxtaposed E- and GC-boxes during early spermatogenesis and oogenesis. PMID: 25003626
- Results identify Sp1 as an inhibitor of DsbA-L gene transcription, and the Sp1-mediated inhibition of DsbA-L gene expression may provide a mechanism underlying obesity-induced adiponectin downregulation and insulin resistance. PMID: 25024375
- Lipopolysaccharide decreases SIGIRR expression by suppressing specificity protein 1 Sp1 via the TLR4-p38 pathway in monocytes and neutrophils. PMID: 24821721
- Increased expression of colonic Wnt9A through Sp1-mediated transcriptional effects involving arylsulfatase B, chondroitin 4-sulfate, and galectin-3. PMID: 24778176
- Sp1/3 transcription factors trigger Mina expression through additive activity targeted to a cluster of four Sp1/3 binding sites forming the P1 promoter. PMID: 24324617
- Sp1 has a critical role at early stages of hematopoietic specification PMID: 24850855
- these findings indicate that miR-200 family members require Sp1 to drive basal expression and to maintain an epithelial state. PMID: 24627491
- Different SUMO isoforms differentially regulate Sp1 to control ocular lens epithelial cell differentiation. PMID: 24706897
- miR-29b mediated ethanol neurotoxicity through the SP1/RAX/PKR cascade. PMID: 24554719
- Sp1 plays a key role in transcriptional regulation of PPARalpha promoter activity and gene expression. PMID: 24451878
- Hypoxic stress downregulated the expression of SIRT1, primarily at the transcriptional level, by reducing the occupancy of the transcriptional activator Sp1 on the proximal promoter of the SIRT1 gene in a SUMOylation-dependent manner. PMID: 23843607
- results show that nonproductive binding of OGG1 to 8-oxoG in promoter sequences could be an epigenetic mechanism to modulate gene expression for a prompt innate immune response. PMID: 24489103
- This study reported on the cloning and characterization of a proximal promoter region and initiated the analysis of transcription factors that control CaV 3.1 channel expression using the murine Cacna1g gene as a model. PMID: 23868804
- show a novel mechanism that links the Rac2-dependent arrest of chemotaxis to decreased expression of PLD2 through the transcription regulator Sp1 PMID: 23898047
- Sp1 is required for IL-15 induction by both poly(I:C) and respiratory syncytial virus, a response that also requires NFkappaB2 and IKKepsilon. PMID: 23873932
- an interplay between Sp1 and Sp3 as a mechanism involved in the control of Fiat gene expression in osteoblasts. PMID: 23463631
- Oct-6 and Oct-11 contribute to the regulation of loricrin gene transcription via interaction with AP-1 factors and Sp1/Sp3. PMID: 23341029
- CRABP2 promotes myoblast differentiation and is modulated by the transcription factors MyoD and Sp1 in C2C12 cells. PMID: 23383201
- Consensus Sp1 sequences, located in the proximal promoter and in the bone-enhancer, as Osterix binding regions in the Col1a1 promoter in vitro and in vivo, were identified. PMID: 23159876
- Sp1 and possibly GATA1 bind concomitantly to the Ccna1 promoter and suppress its activity in vivo. PMID: 23112860
- Sp1 is a key element in the transcriptional regulation of P2X7 receptor in the nervous system. PMID: 23139414
- A conserved region of human and mouse SYN1 promoters contains cis-sites for the transcriptional activator Sp1 in close proximity to REST binding motifs. PMID: 23250796
- These results show that CTCF can bind to the 40-bp and the 100-bp repeat sequences during Murine gammaherpesvirus 68 latency, whereas binding is reduced upon reactivation. In contrast, Sp1 only bound to the 100-bp repeat after reactivation. PMID: 22706977
- Apolipoprotein E4 is deficient in inducing macrophage ABCA1 expression and stimulating the Sp1 signaling pathway PMID: 22984509
- Data suggest that heme is a dispensable ligand in Rev-erb-alpha (nuclear receptor subfamily 1, group D, member 1) activation of connexin 43 transcription by forming a complex with Sp1. PMID: 23201262
- demonstrated that hypoxia-induced CD147 upregulation was mainly mediated by a combined effect of transcription factors HIF-1 and specificity protein 1 (Sp1) on the activation of CD147 promoter PMID: 22678117
- Sp1 level accumulated strongly in early stage and then declined in late stage, which is important for lung cancer cell proliferation and metastasis during tumorigenesis. PMID: 22158040
顯示更多
收起更多
-
亞細胞定位:Nucleus. Cytoplasm.
-
蛋白家族:Sp1 C2H2-type zinc-finger protein family
-
數據庫鏈接:
Most popular with customers
-
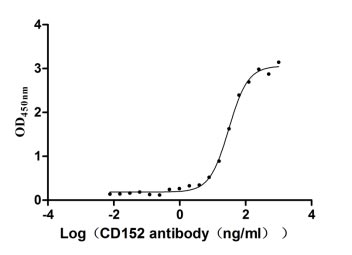
Recombinant Human Cytotoxic T-lymphocyte protein 4 (CTLA4), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
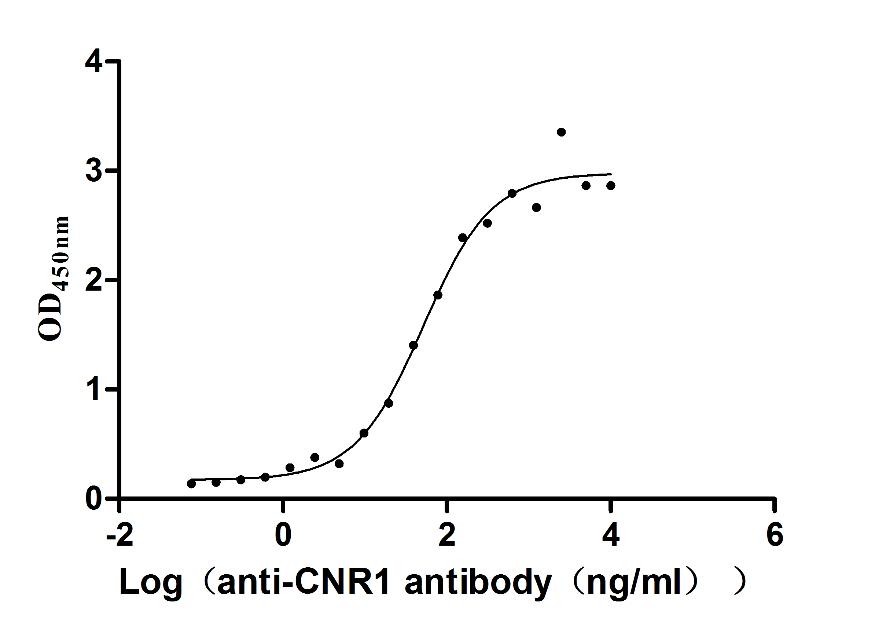
Recombinant Human Cannabinoid receptor 1 (CNR1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
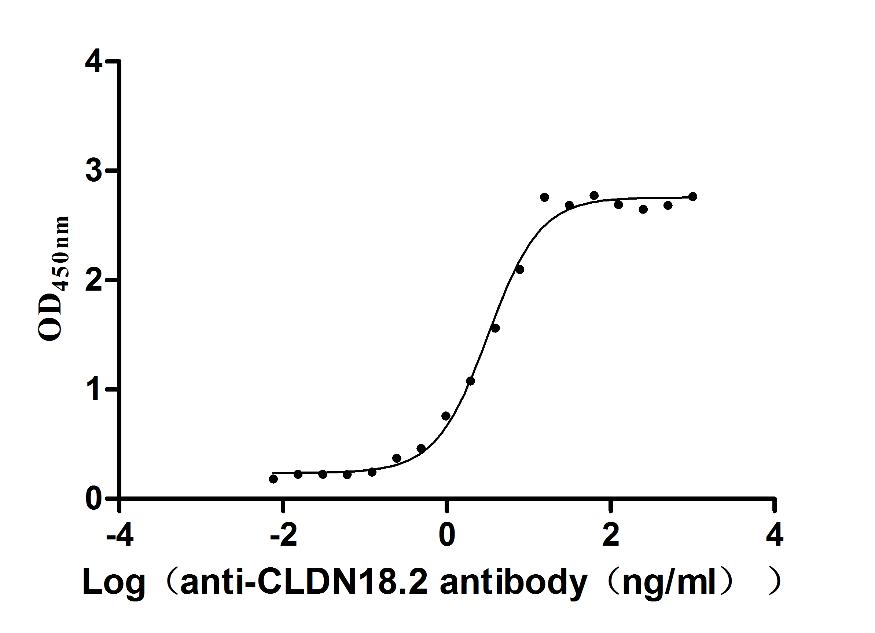
Recombinant Macaca fascicularis Claudin 18.2 (CLDN18.2)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
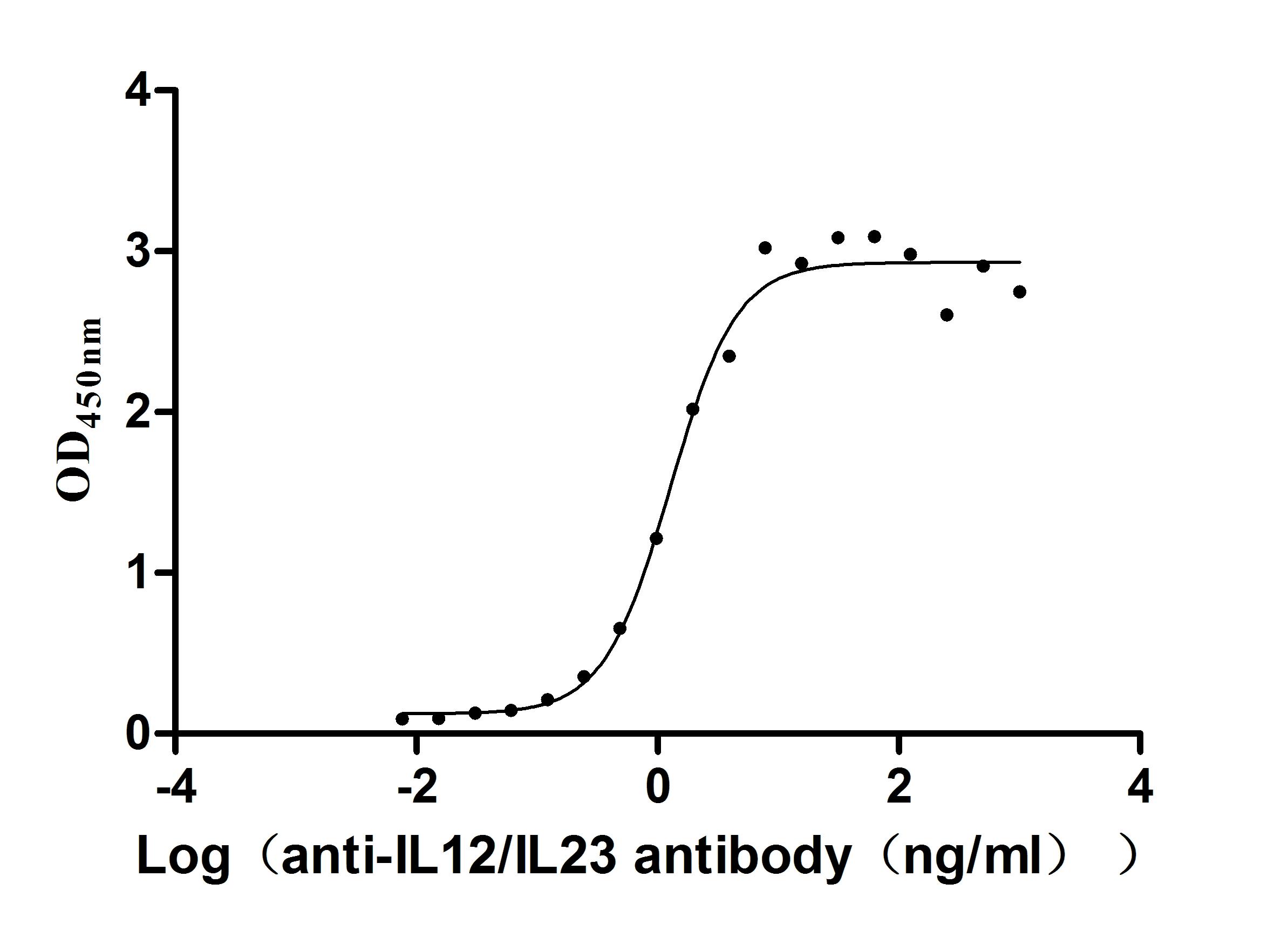
Recombinant Human IL12B&IL12A Heterodimer Protein (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
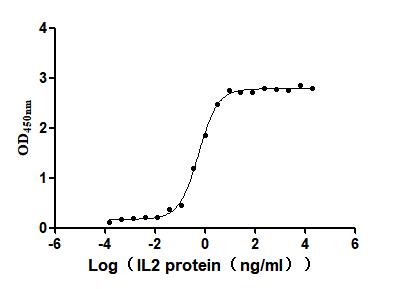
Recombinant Human Interleukin-2 (IL2) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
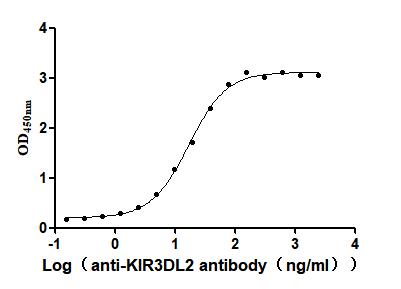
Recombinant Human Killer cell immunoglobulin-like receptor 3DL2 (KIR3DL2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
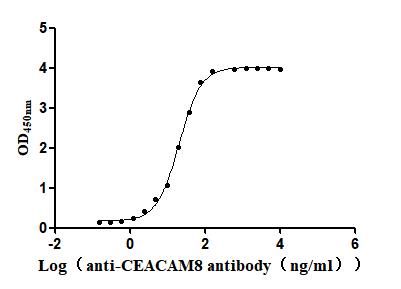
Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 8(CEACAM8) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
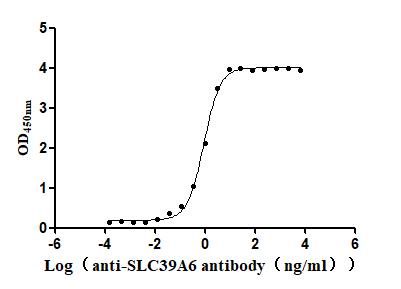
Recombinant Macaca fascicularis Zinc transporter ZIP6 isoform X1(SLC39A6),partial (Active)
Express system: Baculovirus
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)















