-
中文名稱:人SHC1重組蛋白
-
品名簡稱:Recombinant Human SHC1 protein, partial, Biotinylated
-
貨號:CSB-EP021253HU1-B
-
說明書:
-
規格:¥69
-
促銷:
-
圖片:
-
 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產品詳情
-
純度:Greater than 85% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
種屬:Homo sapiens (Human)
-
蛋白長度:Partial
-
來源:E.coli
-
分子量:66.4 kDa
-
表達區域:150-320aa
-
氨基酸序列HPNDKVMGPGVSYLVRYMGCVEVLQSMRALDFNTRTQVTREAISLVCEAVPGAKGATRRRKPCSRPLSSILGRSNLKFAGMPITLTVSTSSLNLMAADCKQIIANHHMQSISFASGGDPDTAEYVAYVAKDPVNQRACHILECPEGLAQDVISTIGQAFELRFKQYLRNPP
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標簽:N-terminal MBP-tagged and C-terminal 6xHis-Avi-tagged
-
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol. If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose.
-
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20°C/-80°C. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Signaling adapter that couples activated growth factor receptors to signaling pathways. Participates in a signaling cascade initiated by activated KIT and KITLG/SCF. Isoform p46Shc and isoform p52Shc, once phosphorylated, couple activated receptor tyrosine kinases to Ras via the recruitment of the GRB2/SOS complex and are implicated in the cytoplasmic propagation of mitogenic signals. Isoform p46Shc and isoform p52Shc may thus function as initiators of the Ras signaling cascade in various non-neuronal systems. Isoform p66Shc does not mediate Ras activation, but is involved in signal transduction pathways that regulate the cellular response to oxidative stress and life span. Isoform p66Shc acts as a downstream target of the tumor suppressor p53 and is indispensable for the ability of stress-activated p53 to induce elevation of intracellular oxidants, cytochrome c release and apoptosis. The expression of isoform p66Shc has been correlated with life span. Participates in signaling downstream of the angiopoietin receptor TEK/TIE2, and plays a role in the regulation of endothelial cell migration and sprouting angiogenesis.
-
基因功能參考文獻:
- Data suggest that up-regulation of SHC threonine phosphorylation is responsible for elevated Akt-signaling and Erk-signaling in triple-negative breast cancer cell lines. PMID: 29208567
- Characterization of bioenergetic parameters and reactive oxygen species production showed that the cellular model of Leigh syndrome is described by increased intracellular oxidative stress and oxidative damage to DNA and proteins, which correlate with increased p66Shc phosphorylation at Ser36. PMID: 28739512
- A positive relationship between the p66Shc expression and oxidative stress was found. p66Shc and oxidative stress were significant predictors of the degree of tubular damage. PMID: 27377870
- Adeno-X Adenoviral System 3 can be used to efficiently construct recombinant adenovirus containing p66Shc gene, and the Adeno-X can inhibit the proliferation of MCF-7 cells by inducing cell cycle arrest at the G2/M phase PMID: 27530145
- STAT4 is a novel transcriptional regulator of p66Shc in normal and chronic lymphocytic leukemia B cells PMID: 27494881
- Isoform b of DDR1 is responsible for collagen I-induced up-regulation of N-cadherin and tyrosine 513 of DDR1b is necessary. PMID: 27605668
- NIC exacerbated AZA-dependent injury via augmenting p66shc transcription. While RES suppressed NIC+AZA-mediated injury, -surprisingly-it further enhanced activity of the p66shc promoter. RES protected cells via the cytoplasmic p66shc/Nrf2/heme oxygenase-1 (HO-1) axis PMID: 28739690
- The results show that the interaction between STS-1 and ShcA is regulated in response to EGF receptor activation. PMID: 28690151
- Nox4-derived H2O2 in part activates Nox2 to increase mitochondrial ROS via pSer36-p66Shc, thereby enhancing VEGFR2 signaling and angiogenesis in endothelial cells. PMID: 28424170
- Data identify, for the first time, a novel non-canonical dynamic mode of interaction between Met and the p66 protein isoform of Shc and its effects on rewiring binding effector complexes according to the activation state of the receptor. PMID: 27048591
- regulates the alternative splicing of XAF1 in extracellular matrix-detachment induced autophagy to coordinate with the anoikic cell death PMID: 26643258
- The silence of p66(Shc) in HCT8 cells reduced the proliferation and accelerated the apoptosis, in addition, the expression of pro-apoptotic proteins caspase-3, caspase-9, Bax was enhanced and the expression of anti-apoptotic protein Bcl-2 was declined. PMID: 26464652
- In mice and humans, reduced p66Shc levels protect from obesity, but not from ectopic fat accumulation, glucose intolerance and insulin resistance. PMID: 26122877
- Data suggest SHC1 (SH2 domain protein C1) expression down-regulates epithelial-mesenchymal transition by repressing TGFB-induced SMAD2/3 activation through differential partitioning of receptors at cell surface of mammocytes/keratinocytes. PMID: 26680585
- p66shc expression in coronary heart disease patients was significantly higher compared with the control group PMID: 24676406
- Finally, a crystal structure of EGFR in complex with a primed Shc1 peptide reveals the structural basis for EGFR substrate specificity. PMID: 26551075
- p53-dependent augmentation of p66(Shc) expression and function represents a key signalling response contributing to beta cell apoptosis under conditions of lipotoxicity PMID: 25810038
- Results show elevated level of p66Shc protein reveal in ovarian cancer cells (OCa) indicating a functional role of the protein in regulating the proliferation of OCa cells. PMID: 24395385
- p66ShcA was upregulated in hearts of patients with ischemic heart disease without heart failure PMID: 25680868
- These data demonstrate that the p52Shc phosphorylation level is altered by the solution environment without affecting the fraction of active c-Src. PMID: 25961473
- Although H2S failed to affect the activities of these two proteins, it disrupted their association. Cysteine-59 resides in proximity to serine-36, the phosphorylation site of p66Shc. PMID: 24766279
- demonstrate that ShcA-dependent activation of AKT, but not the RAS/MAPK pathway, induces VEGF production by bolstering VEGF mRNA translation PMID: 24837366
- the p66shc-dependant ROS production during oxidative stress has mitochondrial origin in human normal and cancer cells. PMID: 24618848
- p66Shc is a bifunctional protein involved in cellular oxidative stress response and differentiation. PMID: 24807908
- High p66Shc expression is associated with malignant gastrointestinal lesions. PMID: 24599562
- CRIF1 knockdown partially induces endothelial activation via increased ROS production and phosphorylation of p66shc PMID: 24906005
- p66(Shc) plays a vital part in canonical Wnt signaling in the endothelium and mediates Wnt3a-stimulated endothelial oxidative stress and dysfunction. PMID: 25147340
- these results identify Grb2 and Shc as central signaling effectors of Met-driven progression of intestinal epithelial-derived cancers. Notably, they suggest that Grb2 may represent a promising target for the design of novel colorectal cancer therapies. PMID: 24708867
- repression of Shc expression by let-7a delays senescence of human diploid fibroblasts. PMID: 24165399
- Study identifies p66ShcA as one of the first prognostic biomarkers for the identification of more aggressive tumors with mesenchymal properties, regardless of molecular subtype. PMID: 25071152
- Serine phosphorylation of p66shc is carried out by active MKK6.beta-Amyloid-induced ROS production and apoptosis increased in the presence of MKK6 and p66shc, which directly associate. PMID: 24085465
- unlike the other isoforms of Shc1, p66Shc appears to antagonize insulin and mTOR signaling, which limits glucose uptake and metabolism. PMID: 24550542
- Exposure of human aortic endothelial cells to stretch led to stretch- and time-dependent p66(Shc) phosphorylation downstream of integrin alpha5beta1 and JNK kinase. In parallel, NADP oxidase and reactive oxygen species increased, and NO bioavailability decreased PMID: 24842918
- study concludes hypercholesterolemia stimulates p66Shc expression in platelets, promoting platelet oxidative stress, hyperreactivity and hyperaggregation via p66Shc PMID: 24845561
- In lung cancer tissues and single cells, p66(Shc) expression inversely correlates with that of Aiolos. PMID: 24823637
- Arg-II promotes mitochondrial dysfunction leading to VSMC senescence/apoptosis through complex positive crosstalk among S6K1-JNK, ERK, p66Shc, and p53, contributing to atherosclerotic vulnerability phenotype. PMID: 23832324
- a central role for adaptor proteins p66Shc and Grb2 in the regulation of ARF1 and ARF6 activation in invasive breast cancer cells. PMID: 24407288
- different domains in SOCS5 contribute to two distinct mechanisms for regulation of cytokine and growth factor signaling with JAK1 and SHC-1 PMID: 23990909
- Salvianolic acid A induced SIRT1 plays an anti-apoptotic role in concanavalin A induced hepatitis by inhibiting p66Shc expression. PMID: 23993977
- p66(Shc) is a good candidate molecule to address the mechanisms underlying healthy aging and to be targeted for the development of novel pharmacological tools for the prevention or cure of age-related pathologies. PMID: 23524280
- DNA methylation of the p66Shc promoter was significantly decreased in the intrauterine growth restricted compared with the appropriate for gestational age infants groups. PMID: 23529764
- Shc plays a pivotal role in coordinately regulating autophagy process and apoptotic resistance in lung adenocarcinoma cells under nutrient-limited conditions. PMID: 23815759
- Knock-down of p66(Shc) leads to a positive feedback upregulation of Nrf2 expression and accordingly, Nrf2 is found to be highly expressed in tumors with low p66(Shc) expression PMID: 23689140
- The p66Shc protein (SHC1 protein) is very important for the regulation of the intracellular redox balance and oxidative stress levels. PMID: 23606925
- Distinct phosphotyrosine-dependent functions of the ShcA adaptor protein are required for transforming growth factor beta (TGFbeta)-induced breast cancer cell migration, invasion, and metastasis PMID: 23277357
- our analysis of human samples demonstrated that enhanced p53/p66Shc signaling plays an important role in the progression of human non-alcoholic fatty liver disease. PMID: 22641095
- Expression of SHC1 mutant protects PC-3 cells against Diallyl trisulfide induced death. PMID: 22020565
- S1P1 expression is controlled by the pro-oxidant activity of p66Shc and is impaired in B-CLL patients with unfavorable prognosis. PMID: 23033271
- Elevated p66Shc expression enhances prostate cancer tumorigenicity. PMID: 22561705
- These findings indicate that LDL cholesterol upregulates human endothelial p66shc expression via hypomethylation of CpG dinucleotides in the p66shc promoter. PMID: 22661506
顯示更多
收起更多
-
亞細胞定位:Cytoplasm.; [Isoform p46Shc]: Mitochondrion matrix.
-
組織特異性:Widely expressed. Expressed in neural stem cells but absent in mature neurons.
-
數據庫鏈接:
Most popular with customers
-

Recombinant Rat Intestinal-type alkaline phosphatase 1 (Alpi) (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
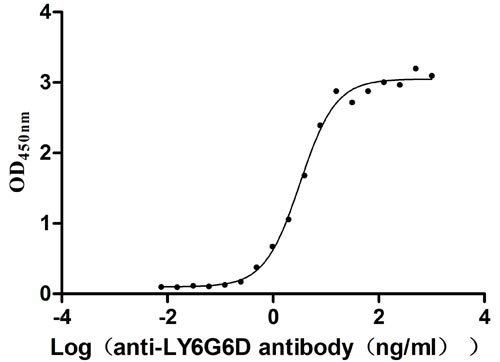
Recombinant Human Lymphocyte antigen 6 complex locus protein G6d (LY6G6D) (Active)
Express system: Yeast
Species: Homo sapiens (Human)
-
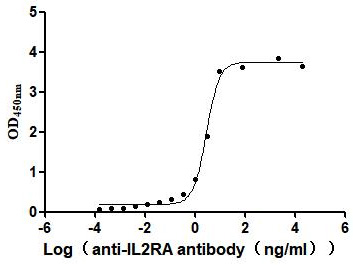
Recombinant Human Interleukin-2 receptor subunit alpha (IL2RA), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
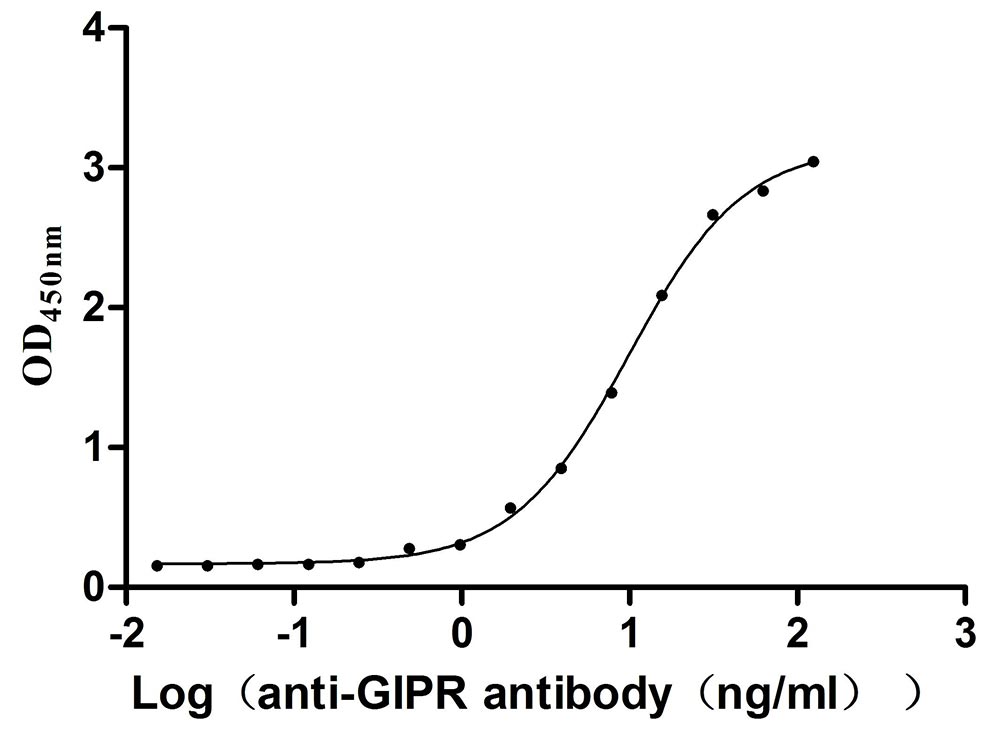
Recombinant Mouse Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
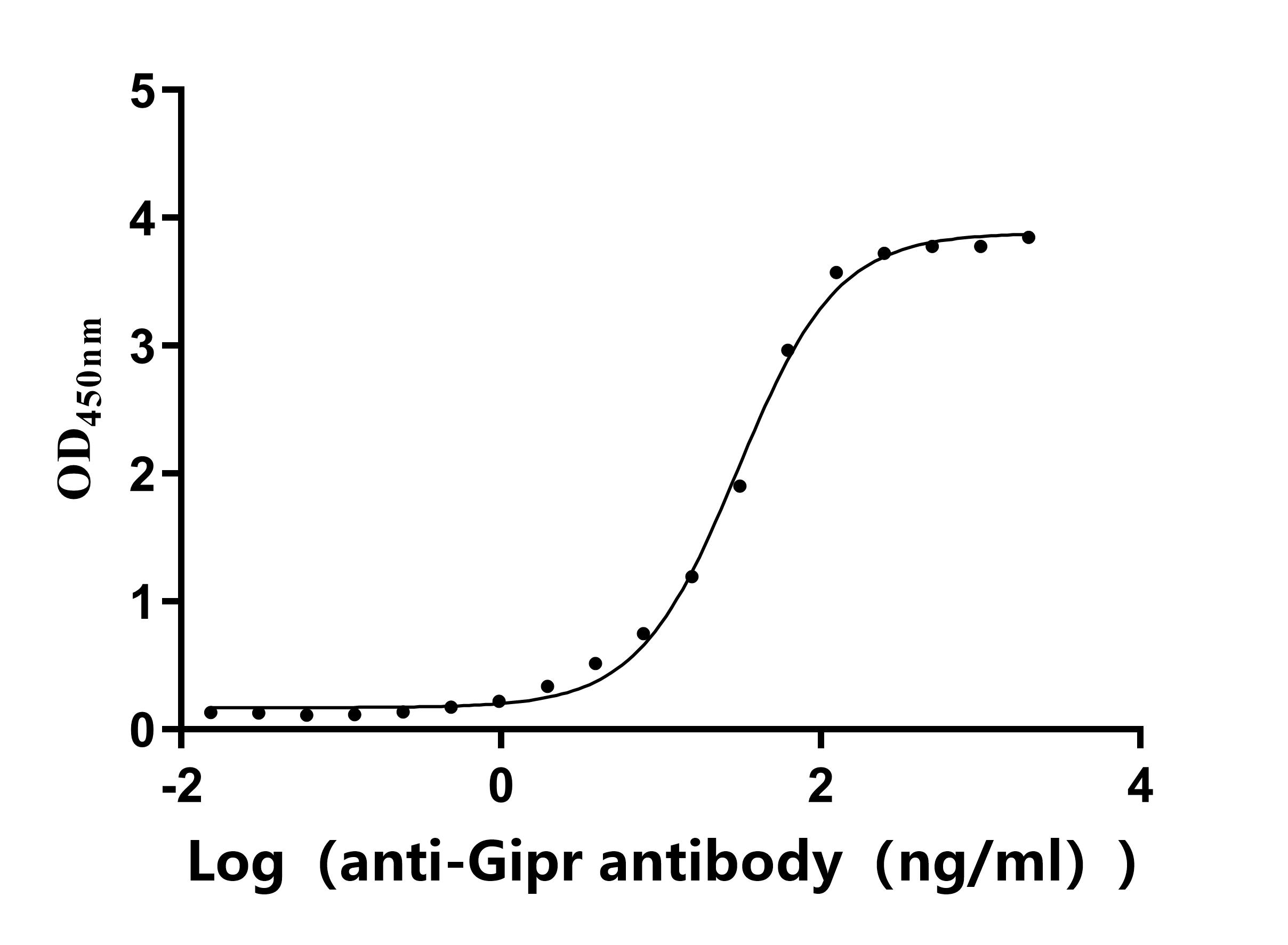
Recombinant Rat Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
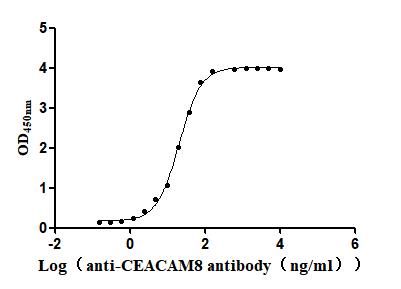
Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 8(CEACAM8) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
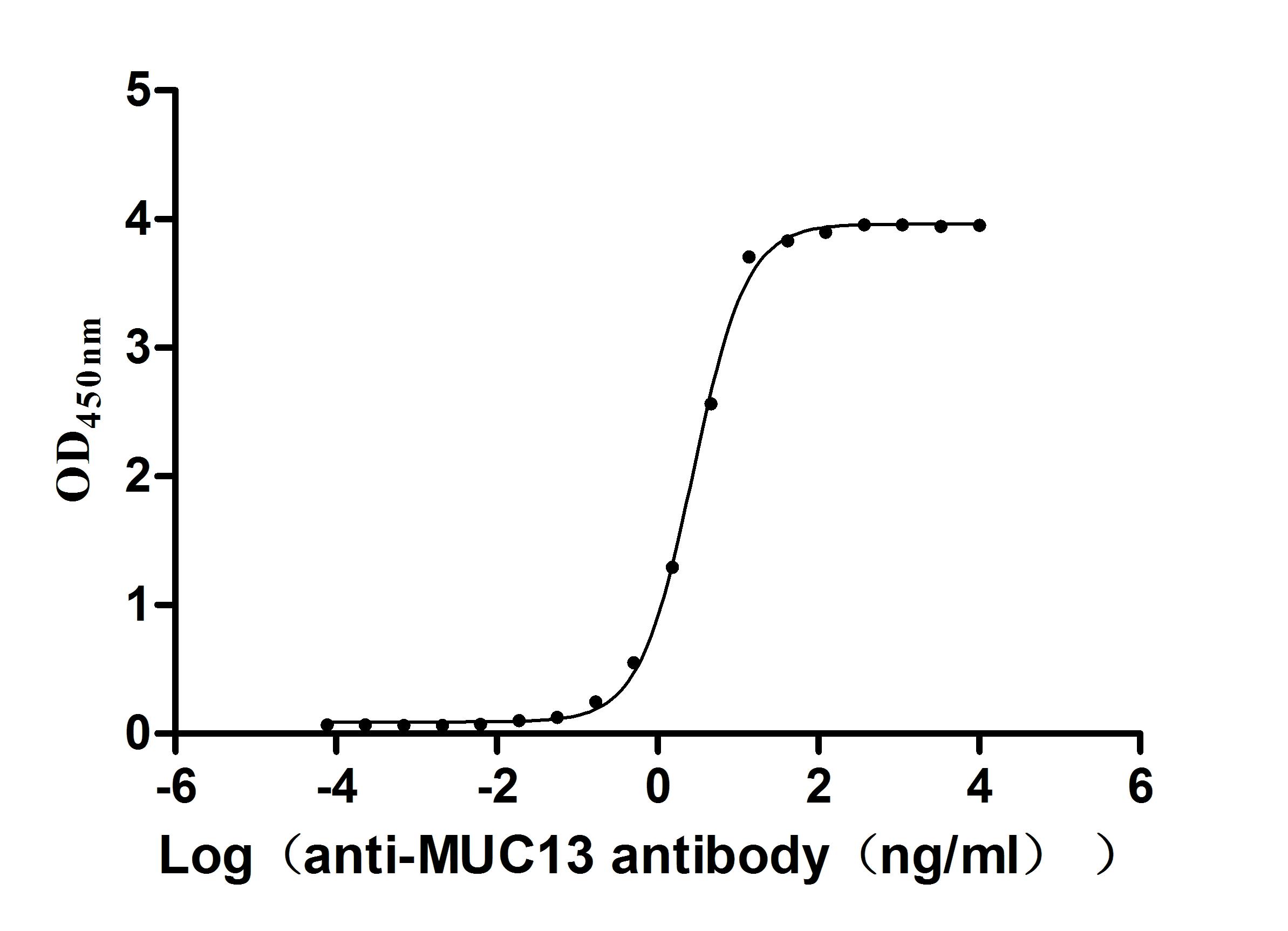
Recombinant Human Mucin-13(MUC13),partial (Active)
Express system: yeast
Species: Homo sapiens (Human)
-
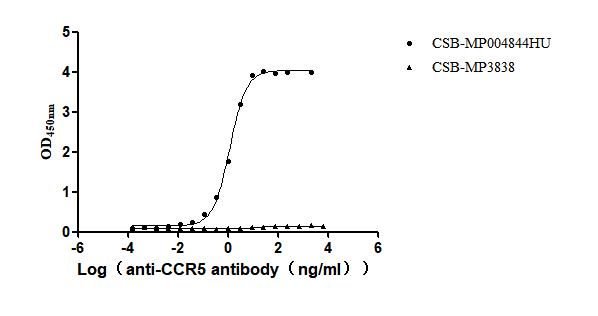
Recombinant Human C-C chemokine receptor type 5 (CCR5)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)















